La régulation de la glycémie
Vidéo en bas de page ^^
I Découverte du rôle du pancréas
Il a fallu un siècle pour éclaircir le système de régulation de la glycémie.
En 1869, Paul Langerhans, anatomo-pathologiste allemand, remarque que le pancréas est constitué de deux catégories de cellules : au milieu des cellules sécrétrices de suc pancréatique, il existe des petits amas de cellules non connectées aux canaux collecteurs du suc pancréatique. Ces amas cellulaires portent aujourd’hui son nom : les îlots de Langerhans. On sait aujourd’hui que ces îlots ont une fonction endocrine, c'est-à-dire qu’ils libèrent des hormones. L’élucidation de leur rôle pris un demi-siècle.
Document 1 : Paul Langerhans, anatomo-pathologiste allemand (1847-1888)

709px-Paul_Langerhans_1878, domaine publique, https://commons.wikimedia.org/wiki/File:Paul_Langerhans_1878.jpg
Document 2 : Coupe de pancréas montrant les îlots de Langerhans
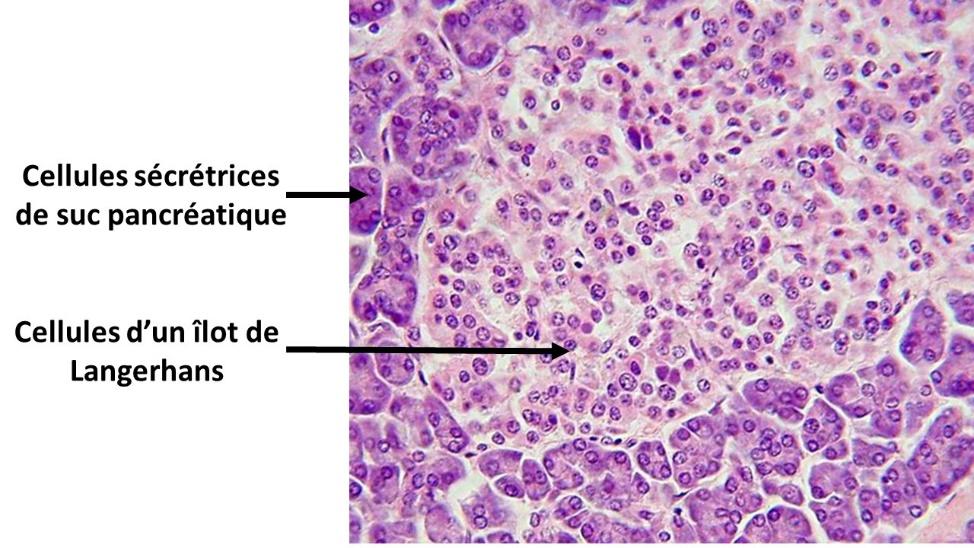
Langerhanssche_Insel, par Polarlys,propre travail, via Wikimédia Commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Langerhanssche_Insel.jpg
Document 3 : Organisation du pancréas
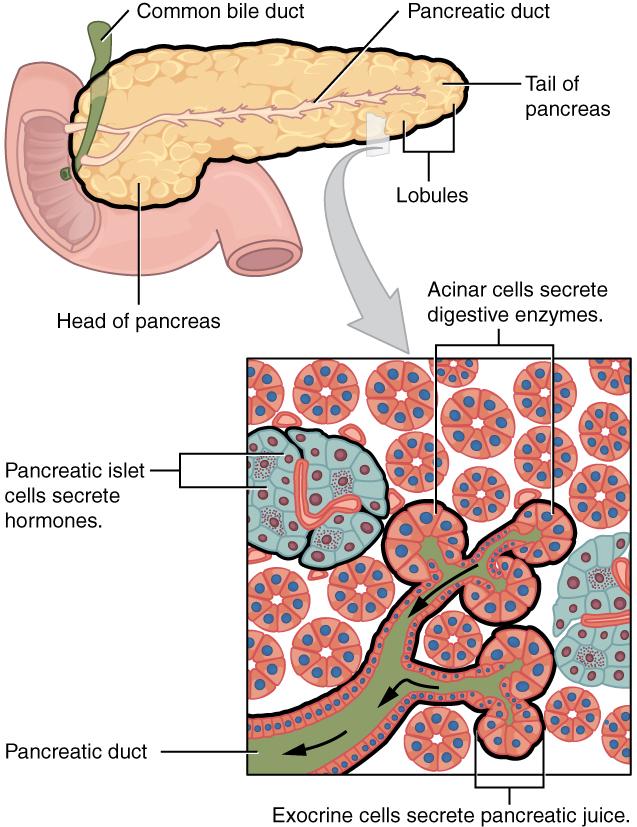
2424_Exocrine_and_Endocrine_Pancreas, par Collège OpenStax, Via wikimedia commons, CC-BY-3.0, https://commons.wikimedia.org/wiki/File:2424_Exocrine_and_Endocrine_Pancreas.jpg
En 1889, Oskar Minkowski et Joseph Von Mering, médecins respectivement allemand et prussien, découvrent que l’ablation du pancréas chez un chien provoque une hyperglycémie. Une ligature du canal de Wirsung transportant le suc pancréatique ne reproduit pas cet effet. Si le pancréas possède donc bien une partie exocrine libérant des sucs digestifs qui n’interviennent pas dans la régulation de la glycémie, il doit donc, en plus de ce rôle, libérer une substance qui régule la glycémie.
Document 4 : Oskar Minkowski (1858-1931) et Joseph Von Mering (1849_1908)


Oskar_Minkowski_(2), https://commons.wikimedia.org/wiki/File:Oskar_Minkowski_(2).jpg
Josef_von_Mering_2, domaine publique, https://commons.wikimedia.org/wiki/File:Josef_von_Mering_2.jpg
Document 5 : Résultats de l’expérience Minkowski et Von Mering

En 1893, le professeur français Emmanuel Hédon, réalisa une expérience de greffe de pancréas chez le chien. Quelques heures après l’ablation du pancréas, il réalisa une greffe de celui-ci au niveau du cou et rétablit la circulation sanguine entre l’animal et le greffon pendant quelques heures avant de l’interrompre. Il remarqua qu’après la greffe, la glycémie baissait puis augmentait de nouveau, une fois le greffon supprimé. Il supposa donc que la régulation de la glycémie par le pancréas se faisait par l’intermédiaire de la régulation sanguine et donc de manière hormonale.
Document 6 : Hédon Emmanuel, professeur de physiologie, Faculté de Médecine de Montpellier (1863-1933)

603px-Professeur_Hédon_CIPH0218, Banque d'images de la Bibliothèque interuniversitaire de santé: CIPH0218, Éléments avec l'autorisation OTRS confirmés, via Wikimédia Commons, https://commons.wikimedia.org/wiki/File:Professeur_H%C3%A9don_CIPH0218.jpg
Document 7 : Résultats de l’expérience du professeur Hédon (1893)

C’est en 1921 que Frédrick Banting (1891-1941) médecin canadien et un jeune médecin américano-canadien, Charles Herbert Best, testent cette hypothèse, et cherchent à isoler l’hormone pancréatique. Pour ce faire, ils enlèvent le pancréas d’un chien puis réalisent diverses injections intraveineuses. Ils injectent tout d’abord de l’extrait de pancréas et constatent une baisse de la glycémie : ils en déduisent que le pancréas contient une substance hypoglycémiante. En injectant des extraits de pancréas bouilli, la glycémie ne baisse pas. Ils en déduisent que le pancréas contient bien une substance hypoglycémiante qui, si elle est dénaturée par la chaleur, n’agit pas. En collaboration avec Macleod (1876-1935) scientifique britannique et avec le chimiste James Collip, ils isolèrent l’hormone en 1921 et soignèrent pour la première fois un jeune patient âgé de 14 ans, Léonard Thompson, atteint de diabète « sucré », lui sauvant au passage la vie.
Document 8 : Frédrick Banting (1891-1941) à droite et Charles Herbert Best (1899- 1978)

C._H._Best_and_F._G._Banting_ca._1924 Bibliothèque et Archives du Canada - PA-123481
via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Fredrick_banting.jpg
Document 9 : John James Richard Macleod ( 1876-1935)

J.J.R._Macleod_ca._1928, source université de Toronto, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:J.J.R._Macleod_ca._1928.png
Document 10 : Résultats de l’expérience de Banting et Best en 1921

L’hormone est d’abord nommée « pancréine » par le médecin roumain Nicolae Paulescu qui avait essayé de l’isoler un an avant Banting mais sans obtenir d’extraits stables. Elle fut ensuite nommée « insuline » par Edward Albert Sharpey-Schafer, physiologiste anglais.
Document 11 : Nicolae Paulescu, médecin roumain (1869-1931)

Nicolae_Paulescu, source http://www.crestinortodox.ro/religie/nicolae-constantin-paulescu-125700.html via wikimedia commons, domaine publique, https://commons.wikimedia.org/wiki/File:Nicolae_Paulescu.jpg
Document 12 : Edward Albert Sharpey-Schafer, physiologiste anglais (1850-1935)

Edward_Albert_Sharpey-Schafer_2, par Lippiatt via wikimedia commons, CC-BY-4.0, https://commons.wikimedia.org/wiki/File:Edward_Albert_Sharpey-Schafer_2.jpg
En 1943, C.J. Dunn constate que l’injection d’une substance chimique, l’alloxane, provoque les mêmes troubles de la glycémie qu’une ablation du pancréas, c’est à dire une augmentation de la glycémie. Il constate que l’alloxane détruit les îlots de Langerhans. Le lien est fait : ces derniers contiennent donc des cellules capables de produire l’hormone hypoglycémiante qu’est l’insuline. Les îlots de Langerhans ont donc une fonction endocrine.
Document 13 : Expérience de Dunn en 1943

En 1923, Charles Kimball et John Murlin émirent l’hypothèse que s’il existait une hormone hypoglycémiante, il devait forcément en exister une qui avait une action antagoniste à savoir une action hyperglycémiante. En 1953, Alfred Staub isole et détermine la composition chimique de cette hormone, le glucagon, elle aussi produite par le pancréas.
Aujourd’hui, grâce aux procédés d’histochimie utilisant des anticorps marqués dirigés contre l’une ou l’autre des deux hormones, il est possible de localiser précisément le lieu de production de celles-ci dans le pancréas. L’insuline est produite par des cellules situées au centre des îlots de Langerhans et que l’on appelle des cellules β. Le glucagon est produit par des cellules situées à la périphérie des îlots de Langerhans et que l’on appelle des cellules α.
Document 14 : Localisation des cellules productrices des hormones régulatrices de la glycémie dans un îlot de Langerhans : cellules β pour l’insuline en bleu et cellules α pour le glucagon en rouge (révélation par immunocoloration)
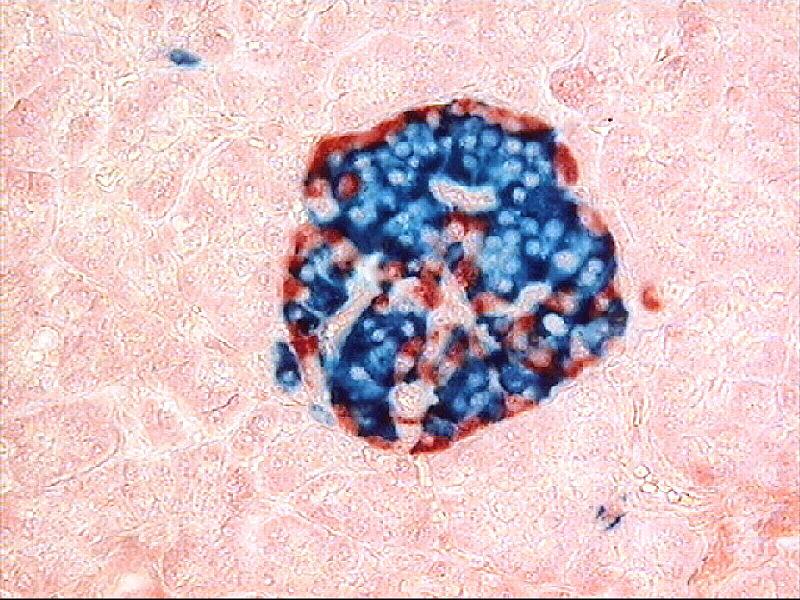
Human_pancreatic_islet, par Afférent propre travail, Généré au Laboratoire de développement du système nerveux, FSBI Human Morphology SRI RAMS, Moscou, via Wikimédia Commons,
II Le rôle du glucagon
Le glucagon est une hormone peptidique sécrétée par les cellules α des îlots de Langerhans. Chez l’Homme, il est composé de 29 acides aminés organisés selon la séquence suivante : NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-
Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr-COOH.
Document 15 : Molécule de glucagon, représentation 3D rubanée

PBB_Protein_GCG_image, par http://en.wikipedia.org/wiki/User:ProteinBoxBot, via Wikimédia communs, domaine publique, https://commons.wikimedia.org/wiki/File:PBB_Protein_GCG_image.jpg
L’injection de glucagon chez un chien à jeun provoque une hausse de la glycémie. On peut donc dire que le glucagon est hyperglycémiant. On peut supposer que la quantité de glucagon libérée dépendra du niveau atteint par la glycémie : plus celle-ci serait basse, plus la quantité de glucagon libérée devrait être importante.
Document 16 : Évolution de la glycémie après injection de glucagon chez un chien à jeun

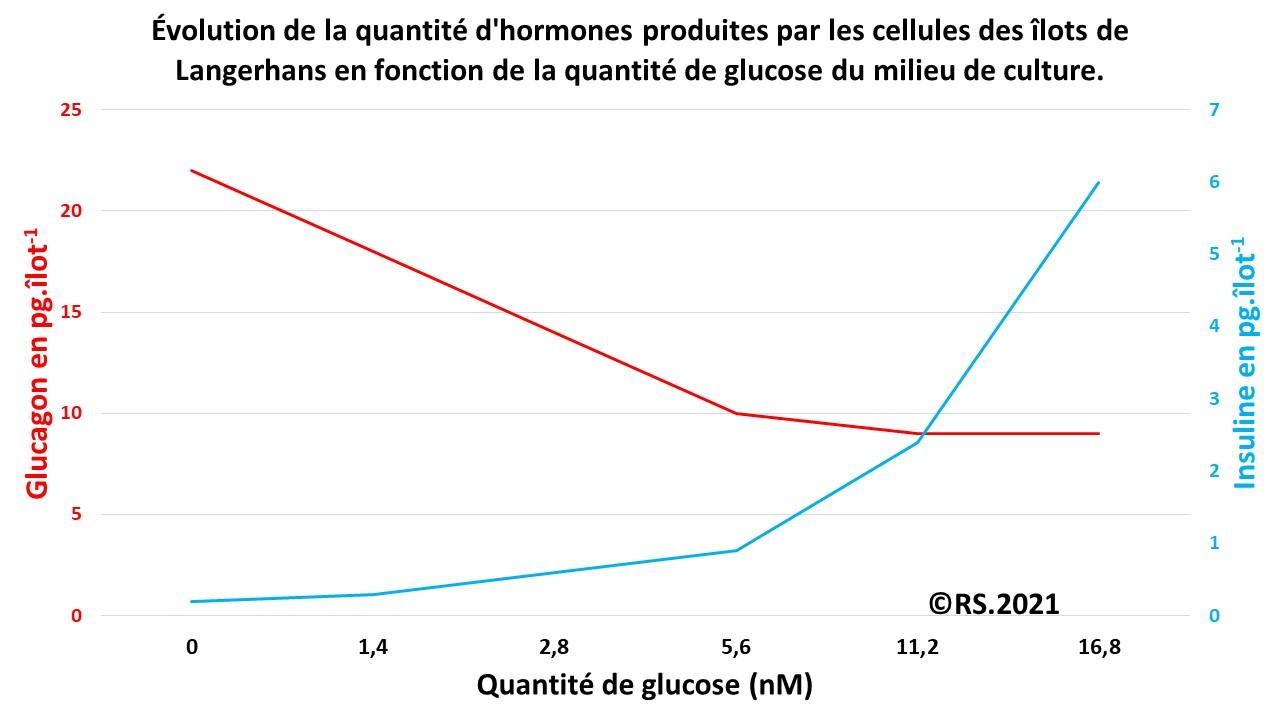
Dans un milieu de culture contenant des îlots de Langerhans, on remarque que plus la quantité de glucose du milieu de culture est importante, plus la concentration en glucagon mesurée dans ce milieu diminue. On en déduit que la quantité de glucagon libérée dépend bien de la concentration en glucose. Le glucagon doit donc être produit en cas de baisse de la glycémie, après des périodes de jeûne ou d’activité physique. On peut supposer qu’il a une action sur les réserves de glucose, facilitant ainsi sa libération
Document 17 : Mesures de la quantité des hormones (glucagon et insuline) produites en fonction de la quantité de glucose du milieu de culture.
L’enregistrement de la teneur en glycogène des cellules hépatiques d’un chien non diabétique à jeun et mis sous perfusion de glucagon confirme cette hypothèse. En effet, après la pose d’une perfusion de 2.5 µg de glucagon pendant 7h, des biopsies régulières du foie révèlent une diminution de la teneur en glycogène cellulaire et on note en parallèle une augmentation de la glycémie du chien. On en déduit que le glycogène du foie est hydrolysé pour libérer du glucose dans le sang.
Document 18 : Évolution de la glycémie d’un chien sous perfusion de glucagon et évolution de la teneur en glycogène de ses cellules hépatiques

On rappelle que le glycogène stocké dans les cellules hépatiques ou musculaires provient de la polymérisation du glucose par une enzyme appelée « glycogène synthase ». Il peut être hydrolysé par une enzyme, la « glycogène phosphorylase ». On peut donc supposer que le glucagon modifie l’activité de ces 2 enzymes.
Document 19 : Équilibre glucose-glycogène

On observe après une perfusion de glucagon, que l’activité de la glycogène synthase diminue et que celle de la glycogène phosphorylase augmente. La première étant responsable de la formation intracellulaire de glycogène à partir du glucose provenant du sang, si son activité diminue alors, la glycémie augmente car le glucose n’est plus stocké sous forme de glycogène. Si l’activité de la glycogène phosphorylase augmente alors la libération de glucose dans le sang est augmentée. La combinaison des 2 actions concourt à augmenter le taux de glucose du sang : le glucagon serait donc une hormone hyperglycémiante modifiant l’activité des enzymes liées au glycogène.
Document 20 : Effet du glucagon sur l’évolution de l’activité des enzymes glycogène synthase et glycogène phosphorylase

On peut se demander comment cette hormone circulant dans le sang, modifie l’activité des enzymes intracellulaires.
Comme toute hormone, le glucagon possède des cellules cibles dont les cellules du foie font partie, en plus de celles du cerveau, des reins et des muscles lisses intestinaux. Il se fixe sur un récepteur membranaire spécifique de type récepteur métabotrope à protéine G, appelé récepteur GPCR. La fixation du glucagon sera à l’origine d’une série de réactions biochimiques modifiant l’activité de la cellule cible. En effet, le glucagon en se fixant sur son récepteur GPCR provoque un changement de conformation de celui-ci à l’origine de la libération de la protéine G grâce à la consommation d’une molécule de GTP. Une des sous unités de celle-ci active l’adénylate cyclase membranaire (AC), qui synthétise l’AMPc ou Adénosine 3’, 5’-monophosphate cyclique. Cette dernière active l’enzyme appelée protéine kinase A (PKA) qui à son tour, active la phosphorylase kinase (PPK), qui elle-même phosphoryle la glycogène phosphorylase b (PYG b), la convertissant en une forme active appelée phosphorylase a (PYG a). La phosphorylase a est l'enzyme responsable de la libération de glucose 1-phosphate à partir de glycogène.
Document 21 : Cascade de réactions suite à la fixation du glucagon sur son récepteur GPCR
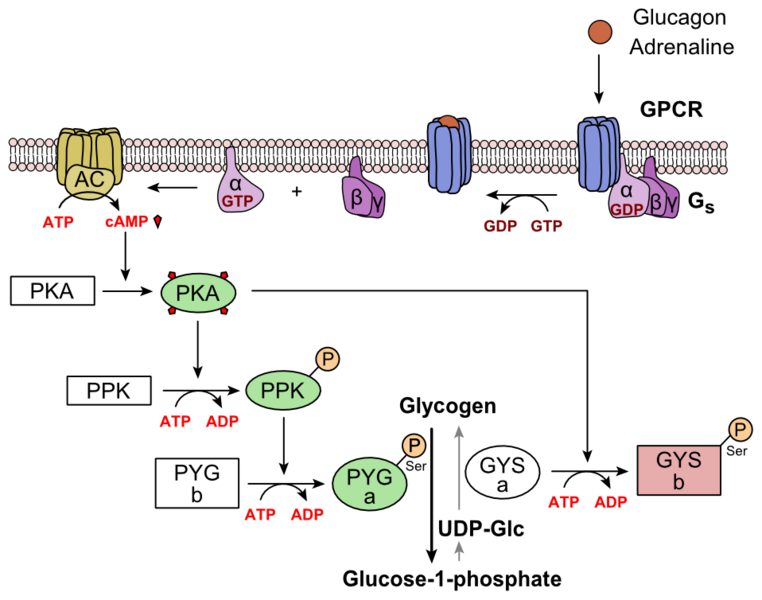
762px-Glucagon_Pathway, par FrozenMan propre travail, via Wikimédia Commons, CC-BY-SA-4.0 https://commons.wikimedia.org/wiki/File:Glucagon_Activation.png
III Le rôle de l’insuline
L’insuline est une hormone peptidique sécrétée par les cellules β des îlots de Langerhans dans le pancréas. C’est une hormone constituée de 2 chaînes polypeptidiques reliées entre elles par 2 ponts disulfures : une chaîne A de 21 acides aminés, et une chaîne B de 30 acides aminés. La structure de l'insuline a été déterminée par Frederick Sanger.
Document 22 : Modèles moléculaires de l’insuline
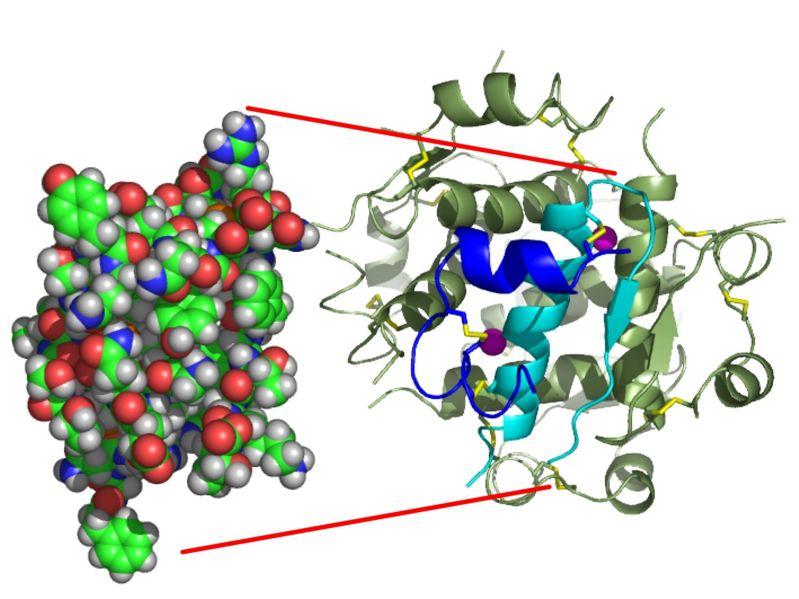
InsulinMonomer, par Isaac Yonemoto., Transféré de en.wikipedia à Commons. Premier téléchargement sur en.wp par Takometer, via Wikimédia Commons, CC-BY-2.5https://commons.wikimedia.org/wiki/File:InsulinMonomer.jpg
Document 23 : Frederick Sanger

675px-Frederick_Sanger2, par Département américain de la santé et des services sociaux . via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Frederick_Sanger2.jpg
Document 24 : îlot pancréatique de souris visualisé par immunofluorescence, taille 326µm
En bleu, les noyaux des cellules voisines, en rouge les anticorps anti-insuline révélant les cellules β de l’îlot de Langerhans.
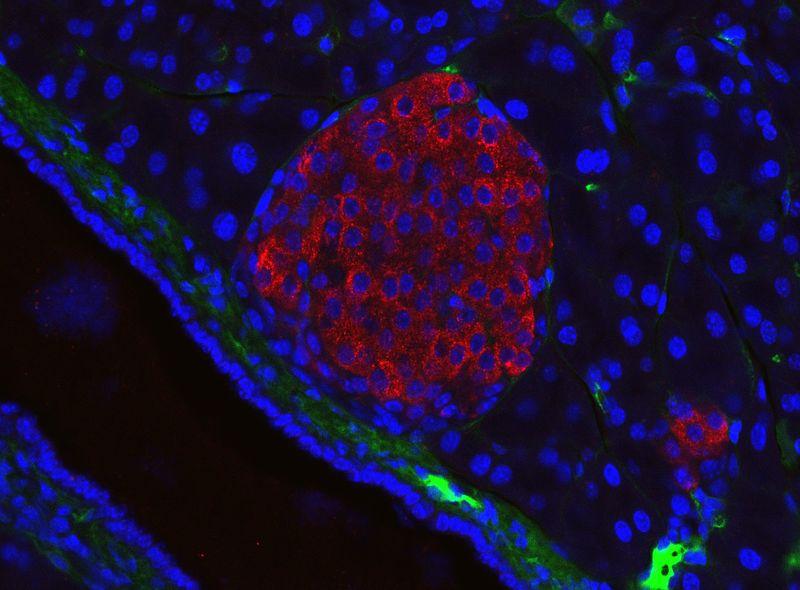
1200px-Mouse_pancreatic_islet, par Jakob Suckale prorpe travail, généré dans le laboratoire Solimena, Institut Paul Langerhans de Dresde, via wikimedia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Mouse_pancreatic_islet.jpg
On réalise des injections d’insuline puis de glucagon à un chien et on mesure la variation de la glycémie. On remarque qu’en cas d’injection d’insuline, la glycémie diminue : l’insuline est donc une hormone hypoglycémiante, c’est à dire une hormone provoquant le stockage du glucose sanguin.
Document 25 : Évolution de la glycémie après injection d’hormones pancréatiques

On peut donc supposer que la quantité d’insuline libérée dépendra de la valeur de la glycémie. Or cette dernière dépend de notre alimentation.
Tous les aliments ne contiennent pas la même quantité de glucose. L'indice glycémique est un critère de classement des aliments contenant des glucides, basé sur leurs effets sur la glycémie durant les deux heures suivant leur ingestion. Il donne une indication sur la capacité d'un aliment à augmenter la glycémie, c'est-à-dire la concentration de glucose dans le sang. L'index glycémique d'un aliment se mesure par rapport à celui du glucose qui est de 100. Un index glycémique de 80 signifie que l'augmentation de la glycémie avec cet aliment correspond à 80 % de celle obtenue avec du glucose.
Document 26 : Indice glycémique de certains aliments
|
|
|
|
|
|
|
|
|
|
|
|
|
|
On enregistre la variation de l’insulinémie (taux d’insuline dans le sang) après ingestion d’aliments. On remarque que la sécrétion d’insuline est différente en fonction des aliments : elle est d’autant plus forte que l’aliment possède un indice glycémique élevé.
Document 27 : Sécrétion d’insuline déclenchée par l’ingestion des aliments précédents
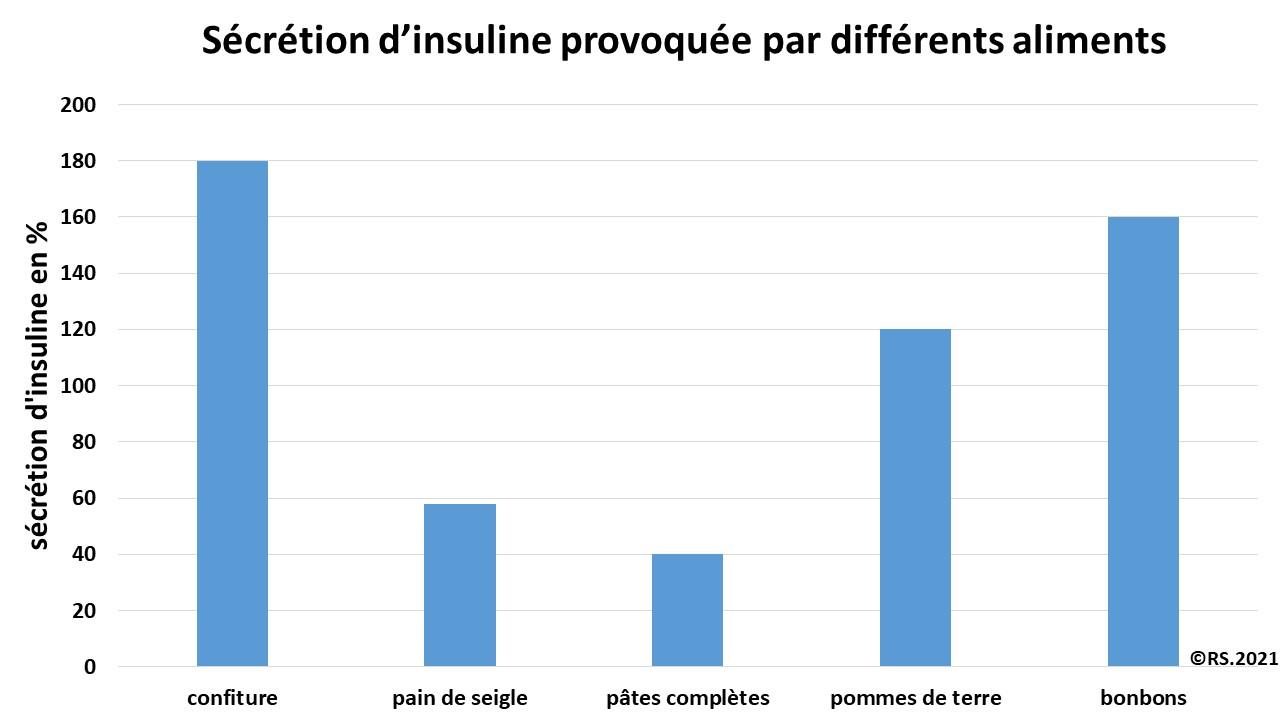
Quel que soit l’apport glycémique, le stockage du glucose permet de maintenir la glycémie. Ainsi en cas d’apport en glucose, une décharge d’insuline permettrait le stockage de celui-ci sous forme de glycogène.
Chez un chien pancréatectomisé, des biopsies régulières montrent une diminution constante de la teneur en glycogène du foie. Des injections d’insuline permettent au foie de retrouver un stock de glycogène élevé.
Document 28 : Effet de l’insuline sur le glycogène hépatique chez un chien pancréatectomisé

On peut donc supposer que dans les cellules hépatiques, l’insuline augmente l’activité de l’enzyme responsable de la glycogénogenèse, la glycogène synthase, et diminue celle de la glycogène phosphorylase.
L’enregistrement de l’activité de ces dernières en fonction de la quantité d’insuline le confirme. Plus il y a d’insuline dans le milieu, plus la glycogène synthase est active et moins la glycogène phosphorylase l’est.
Document 29 : Effet de l’insuline sur l’activité des enzymes liées au glycogène

Ainsi, la mise en réserve du glucose sous forme de glycogène, dans le foie se déroule comme suit : l’insuline stimule la glycogénogenèse en augmentant l’activité de la glycogène synthase et en diminuant celle de la glycogène phosphorylase.
Ceci se vérifie aussi très bien dans le muscle. On perfuse un muscle avec des doses croissantes d’insuline et on mesure le flux de glucose entrant dans ce dernier. La valeur 100 correspond au flux maximum mesurable. On remarque que plus la quantité d’insuline perfusée est importante plus le flux entrant de glucose dans la cellule musculaire est important.
Document 30 : Évolution du flux entrant de glucose dans un muscle sous perfusion d’insuline

Ainsi l’insuline favorise l’absorption et l’utilisation du glucose par les tissus musculaires.
Au niveau des cellules cibles, l’insuline facilite la pénétration du glucose dans les cellules en augmentant la perméabilité au glucose de leur membrane, grâce à la stimulation de la synthèse d’un récepteur au glucose GLUT4 et à la translocation de celui-ci vers la membrane.
Document 31 : Effet de la fixation de l’insuline sur son récepteur

L'insuline se lie à son récepteur (1), qui, à son tour, déclenche de nombreuses cascades d'activation des protéines kinases (2). Celles-ci sont responsables de la translocation du transporteur GLUT4 vers la membrane plasmique (3) et l'afflux de glucose (4), la synthèse du glycogène (5), la glycolyse (6) et la synthèse des acides gras (7).
IV Homéostasie glycémique : un système de régulation
On appelle homéostasie glycémique la capacité d'un système à maintenir la glycémie à sa valeur consigne, quelles que soient les contraintes externes.
L’homéostasie est un phénomène par lequel un facteur clé (par exemple, la température) est maintenu autour d'une valeur bénéfique pour le système considéré, grâce à un processus de régulation. Des exemples typiques d'homéostasie sont : la température d'une pièce grâce à un thermostat, la température du corps d'un animal homéotherme, le taux de sucre sanguin, le degré d'acidité d'un milieu, la pression interne d'un milieu... Plus globalement, on désigne aussi par homéostasie la capacité globale d'un système à maintenir tout un ensemble de facteurs clés, notamment chez un organisme vivant. Le concept de l'homéostasie a été initié par le médecin et physiologiste français Claude Bernard en 1866.
La concentration plasmatique en glucose constitue un paramètre biologique réglé à une valeur de 1g.L-1 de plasma : la valeur consigne. Le maintien de cette constante est assuré par l’intervention permanente d’un système réglant qui est constitué de capteurs : les cellules α et β des îlots de Langerhans (pancréas endocrine) qui détectent les écarts de la glycémie par rapport à la valeur consigne. Les îlots sont richement vascularisés et réagissent en sécrétant davantage d’insuline ou davantage de glucagon. La quantité de ces hormones circulant dans le sang a valeur de message. Les effecteurs, dont les cellules possèdent les récepteurs aux hormones, répondent en corrigeant l’écart constaté et ramènent la glycémie à la valeur consigne : le foie en libérant ou en absorbant du glucose, les muscles et les cellules adipeuses principalement en absorbant du glucose.
Document 32 : Homéostasie glycémique

En cas d’hyperglycémie (glycémie située au-dessus de la valeur consigne), les cellules β produisent d’avantage d’insuline, afin d’éviter une élévation de la glycémie au-delà de 3 g.L-1. C’est une hormone hypoglycémiante qui agit sur les cellules cibles spécifiques en se fixant sur des récepteurs membranaires par complémentarité de forme. Pour cela, elle stimule :
- La production de transporteurs de glucose (ce qui augmente la perméabilité de la membrane au glucose), permettant l’entrée de glucose dans les cellules et sa consommation par celles-ci,
- La glycogénogenèse en augmentant l’activité de la glycogène synthase entraînant la polymérisation des molécules de glucose permettant le stockage du glucose sous forme de glycogène, par les cellules hépatiques et musculaires,
- La lipogenèse avec stockage du glucose sous forme de triglycérides (un glycérol associé à 3 acides gras), dans le foie et les adipocytes.
En cas d’hypoglycémie (glycémie située en dessous de la valeur consigne), à la périphérie des îlots de Langerhans, les cellules α produisent le glucagon afin d’éviter une baisse de la glycémie sous 0.5 g.L-1. C’est une hormone hyperglycémiante dont l’action est essentiellement hépatique en diminuant le stockage du glucose sous forme de glycogène :
- il baisse l’activité de la glycogène synthase.
- il augmente l’activité de la glycogène phosphorylase et donc stimule la dégradation (hydrolyse) du glycogène dans le foie, afin de libérer du glucose dans le sang, et donc le mettre à la disposition de tous les organes (hydrolyse du glycogène : glycogénolyse).
La réserve musculaire est dite « égoïste » car il n’y a pas de mise à disposition de cette réserve pour les autres organes. Les muscles n’interviennent donc pas dans la régulation de l’hypoglycémie.
En cas d’effort musculaire prolongé, afin d’éviter une hypoglycémie, quand les ressources en glycogène musculaire et hépatique sont presque épuisées, d’autres voies métaboliques prennent le relais dans l’apport en glucose aux cellules musculaires.
Le tissu adipeux est la réserve la plus importante de l’organisme mais son utilisation est très indirecte. Il faut que les triglycérides, par lipolyse, soient transformés en glycérol et en acides gras. Le glycérol sera transporté par le sang jusqu’au foie pour y être transformé en glucose par néoglucogenèse. Les acides gras intègreront la mitochondrie et subiront une β –oxydation à l’origine de molécules d’acétylCoA qui pourront intégrer le cycle de Krebs.
Document 33 : Origine et utilisation du glucose par la cellule musculaire

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dans ce système homéostasique, il existe donc un paramètre réglé et un système réglant.
Le système réglant comporte :
- Un système de capteurs d’informations détectant les écarts de glycémie et constitué des cellules des îlots de Langerhans
- Un système de messagers hormonaux (insuline et glucagon)
- Un système effecteur permettant de corriger les écarts comprenant le foie, les muscles et le système adipeux.
Régulation de la glycémie - SVT - SANTÉ Term spé #12 - Mathrix
Date de dernière mise à jour : 29/06/2021