Glycolyse et voie aérobie
Vidéo en bas de page ^^
Introduction
L'ATP (adénosine triphosphate) est la molécule utilisée comme carburant par nos cellules pour toutes les réactions chimiques qui nécessitent de l'énergie. Il n'existe aucun stock réel d'ATP dans l'organisme, c'est pourquoi la moindre consommation d'ATP doit être compensée par une production équivalente, de façon à ce que sa concentration soit la plus stable possible.
Document 1 : Cycle de l’ATP

ADP_ATP_cycle par Muessig propre travail, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:ADP_ATP_cycle.png
Lors d’une activité physique, le stock d’ATP doit donc être rapidement renouvelé. On sait qu’en cas de malaise, un carré de sucre permet de rapidement « retrouver ses forces », en d’autres termes de rapidement reconstituer le stock d’ATP. Il existe donc des voies métaboliques, c'est-à-dire des suites de réactions permettant d'aller d'une molécule à une autre, qui sont génératrices d’ATP à partir de glucose.
En pratique sportive, il est courant d’observer des efforts intenses et rapides qui se réalisent sans un apport en dioxygène, l’appareil respiratoire n’ayant pas eu le temps de s’adapter à l’effort. D’autres sports se pratiquent en apnée, d’autres nécessitent par contre une régulation importante de la respiration. On peut donc penser qu’il existe 2 voies principales de production d’ATP : une voie aérobie qui consomme du dioxygène et une voie anaérobie qui ne consomme pas de dioxygène. Dans tous les cas, ces voies consomment du glucose.
Comment l’organisme produit-il de l’ATP à partir du glucose ?
I La Glycolyse, une première utilisation du glucose
La glycolyse correspond à la dégradation du glucose en pyruvate, libérant au passage 2 ATP. Afin de montrer en quoi consiste la glycolyse, une expérience peut être faite.
Des cultures de levures sont réalisées en présence ou en absence de dioxygène dans des milieux contenant une faible quantité de glucose marqué au carbone 14. Des prélèvements sont effectués à différents moments pour permettre de détecter et d’identifier les molécules radioactives présentes dans les compartiments intracellulaires ou extracellulaires.
Document 2 : Résultats de l’expérimentation sur les levures cultivées en milieu enrichi en glucose marqué au carbone 14
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
||
Que ce soit en présence ou en absence de dioxygène, on constate qu’au cours du temps le glucose absent du milieu intracellulaire en début d’expérience (temps T0) passe dans celui-ci (temps T1).
Au bout du temps T2, le glucose est totalement entré dans la cellule car il n’existe plus dans le milieu extracellulaire. Cependant on ne le trouve plus dans le milieu intracellulaire : on trouve du pyruvate.
On peut donc dire que le glucose a été transformé en pyruvate dans le milieu intracellulaire et ce, que ce soit en présence ou en absence de dioxygène.
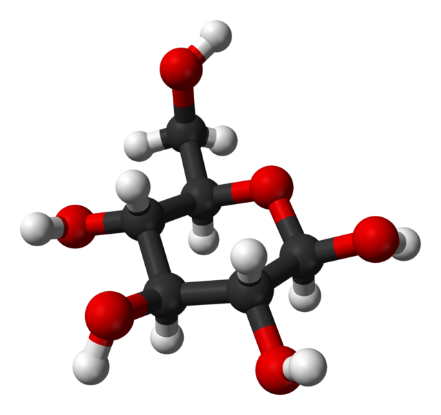
Document 3 : Molécules de glucose et de pyruvate
638px-Beta-D-glucose-from-xtal-3D-balls par Ben Mills propre travail, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Beta-D-glucose-from-xtal-3D-balls.png
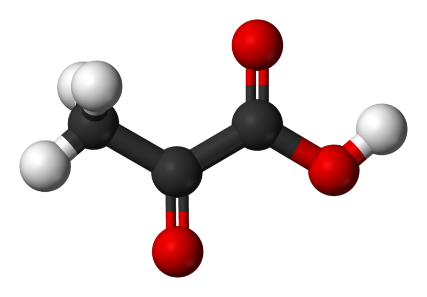
800px-Pyruvic-acid-3D-ballspar Benjah-bmm27 via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Pyruvic-acid-3D-balls.png
En comparant les deux molécules, on constate qu'il y a proportionnellement moins d'hydrogène dans le pyruvate que dans le glucose. Le pyruvate est donc plus oxydé que le glucose. La transformation du glucose en pyruvate est donc une oxydation appelée glycolyse. Elle consiste donc en la transformation de chaque molécule de glucose (à 6 atomes de carbone) en deux molécules d'acide pyruvique (ou pyruvate) à 3 atomes de carbone. Littéralement, la molécule de glucose est « cassée en deux », ce qui produit un peu d'énergie. Au cours de ce processus catalysé par des enzymes, on assiste à des réactions d'oxydoréduction au cours desquelles un accepteur d'électrons (coenzyme R) est réduit.
Cette réduction s’observe très bien expérimentalement grâce au bleu de méthylène. En effet, le bleu de méthylène présente un double intérêt : d'une part, à faible concentration il n'est pas toxique pour les cellules vivantes et, d'autre part, c'est un indicateur d'oxydoréduction. À l'état oxydé il est bleu tandis qu'à l'état réduit il est incolore.
On réalise l’expérience suivante : on prend 2 moules et l’on en fait bouillir une. Chacune des moules est installée dans un tube à essai avec de l’eau de mer puis est recouverte de 3 gouttes de bleu de méthylène peu concentré. On observe le résultat au bout de 2 heures.
Document 4 : Expérience mettant en évidence une réaction d’oxydo-réduction lors de la glycolyse.
T= 0 min T =2 heures
Légende : V = moule vivante, M = moule morte car cuite.


On observe que lorsque les cellules sont vivantes et respirent, le bleu de méthylène s’éclaircit, alors que ce n’est pas le cas lorsque les cellules sont mortes. Or le bleu de méthylène est incolore à l’état réduit. On peut en déduire que la respiration de la moule entraîne la réduction du bleu de méthylène. Il y a donc libération d’électrons qui sont cédés au bleu de méthylène. Ce processus ne se déroule pas dans les cellules mortes et le liquide qui les environne reste bleu, le bleu de méthylène restant à l'état oxydé.
La glycolyse donne donc bien des composés réduits. Il faut donc un accepteur d'électron R qui, en même temps qu’il récupère des électrons, va récupérer des protons (H+) du glucose et deviendra RH2.
R + 2 H+ + 2 e- → RH2
La glycolyse est une oxydation incomplète du glucose, puisqu’il reste à la fin un autre composé organique : le pyruvate.
Glucose + 2 R + 4 H+ + 4 e- → 2 pyruvates + 2 RH2
Cet accepteur d’électrons est le NAD+, c'est-à-dire du nicotinamide adénine dinucléotide sous forme oxydée. Il est réduit en NADH2.
Cette réaction est couplée à la formation de deux molécules d’ATP à partir de deux molécule d’ADP (Adénosine Diphosphate) et de deux phosphates inorganiques notés Pi (phosphate d'hydrogène H3PO4).
2 ADP + 2 Pi → 2 ATP + 2 H2O
Ainsi :
C6H12O6 + 2 ADP + 2 Pi + 2 R → 2C3H4O3 + 2ATP + 2RH2+ 2H2O
Document 5 : Bilan de la glycolyse
©RS.2021

Notons qu’on ne produit seulement que 2 molécules d'ATP pour chaque molécule de glucose : ce bilan est donc relativement faible. Il doit donc exister d’autres réactions qui vont dégrader le pyruvate et produire de l’ATP. Les cellules musculaires produisant de l’ATP en aérobiose ou en anaérobiose, on peut donc penser que le pyruvate, produit dans le cytosol (liquide de base du cytoplasme) lors de la glycolyse sera dégradé à son tour lors d'une seconde étape soit en présence de dioxygène soit en l’absence de celui-ci.
II La respiration cellulaire ou voie aérobie
A/ Localisation de la respiration
La respiration semble se dérouler dans la mitochondrie. En effet, quand des levures sont élevées en milieu aérobie ou en milieu anaérobie, seules les premières possèdent des mitochondries. Cet organite n’est donc présent que lorsque le milieu contient du dioxygène. C’est donc le lieu de la respiration cellulaire.
Document 6 : Comparaison de 2 levures cultivées en présence ou en absence de dioxygène
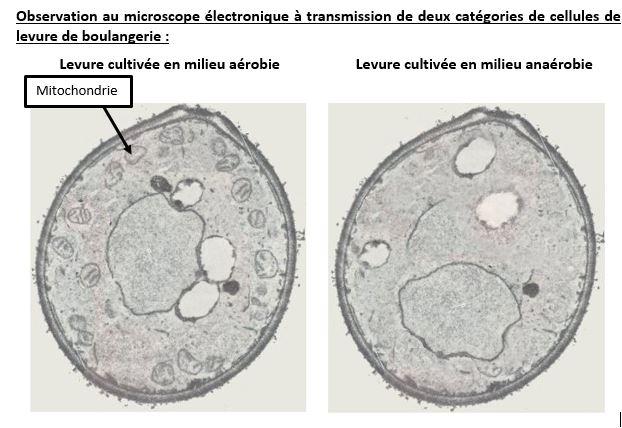
Levures en milieu aérobie et anaérobie, par Alain Gallien,
http://svt.ac-dijon.fr/schemassvt/spip.php?article40
La mitochondrie est un organite présent dans le cytoplasme des cellules eucaryotes Elle proviendrait de l’endosymbiose d’une cellule procaryote. En forme de bâtonnet, elle est remplie d'un liquide, la matrice mitochondriale, et enveloppée par une double membrane, la membrane mitochondriale externe et la membrane mitochondriale interne, qui délimitent un espace intermembranaire.
Chaque mitochondrie possède son propre matériel génétique, indépendant de celui de la cellule qui l'héberge, de sorte qu'elle peut assurer sa reproduction en fonction des besoins de la cellule. Cet ADN est libre dans la matrice. La membrane interne se replie de nombreuses fois à l'intérieur de la matrice pour former des structures très particulière, les crêtes mitochondriales.
Document 7 : Structure d’une mitochondrie

622px-Animal_mitochondrion_diagram_fr.svg, traduit par Ethan Gray , original par LadyofHats, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Animal_mitochondrion_diagram_fr.svg
B/ Les étapes moléculaires de la respiration
1) Mise en évidence du substrat de la respiration mitochondriale
Le glucose est source d’énergie du muscle. Les bactéries utilisent aussi le glucose comme source d’énergie. Cela signifie qu’elles puisent leurs ressources dans le milieu extérieur. Les mitochondries proviendraient de l’endosymbiose d’une archéobactérie. On peut donc se demander si la mitochondrie des cellules eucaryotes utilise directement le glucose qu’elle puiserait dans le hyaloplasme ou si elle utilise le pyruvate issu de la glycolyse.
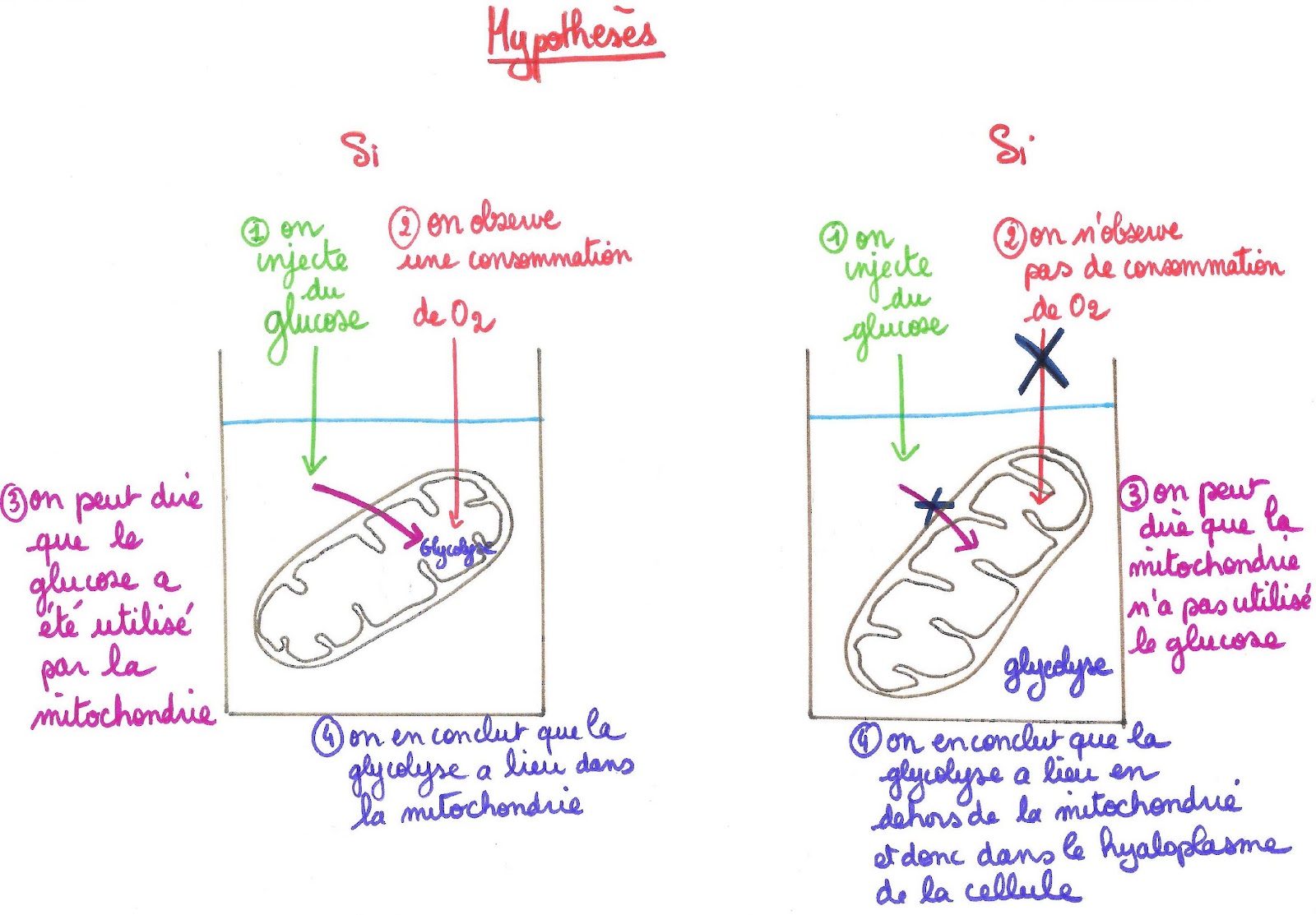
Pour déterminer le rôle des mitochondries, il suffit de tester l’activité respiratoire de celles-ci quand on leur fournit l’une ou l’autre des 2 molécules. L’expérimentation assistée par ordinateur permet d’enregistrer en direct la consommation en dioxygène d’une solution pure de mitochondries grâce à une sonde à dioxygène reliée à une interface connectée à un ordinateur équipé d’un logiciel de mesures.
Si en injectant du glucose dans une solution de mitochondries isolées, on observe une consommation de dioxygène, alors on pourra dire que la mitochondrie consomme le glucose, et qu’une glycolyse a lieu dans la matrice de la mitochondrie. S’il n’y a pas consommation de dioxygène cela signifie que la mitochondrie ne consomme pas de glucose et que la glycolyse n’a pas lieu dans la mitochondrie mais bien dans le hyaloplasme de la cellule eucaryote (elle ne peut avoir lieu dans cette expérience car la mitochondrie baigne dans une solution différente de celle du hyaloplasme).
Document 8 : Hypothèses sous forme de schémas
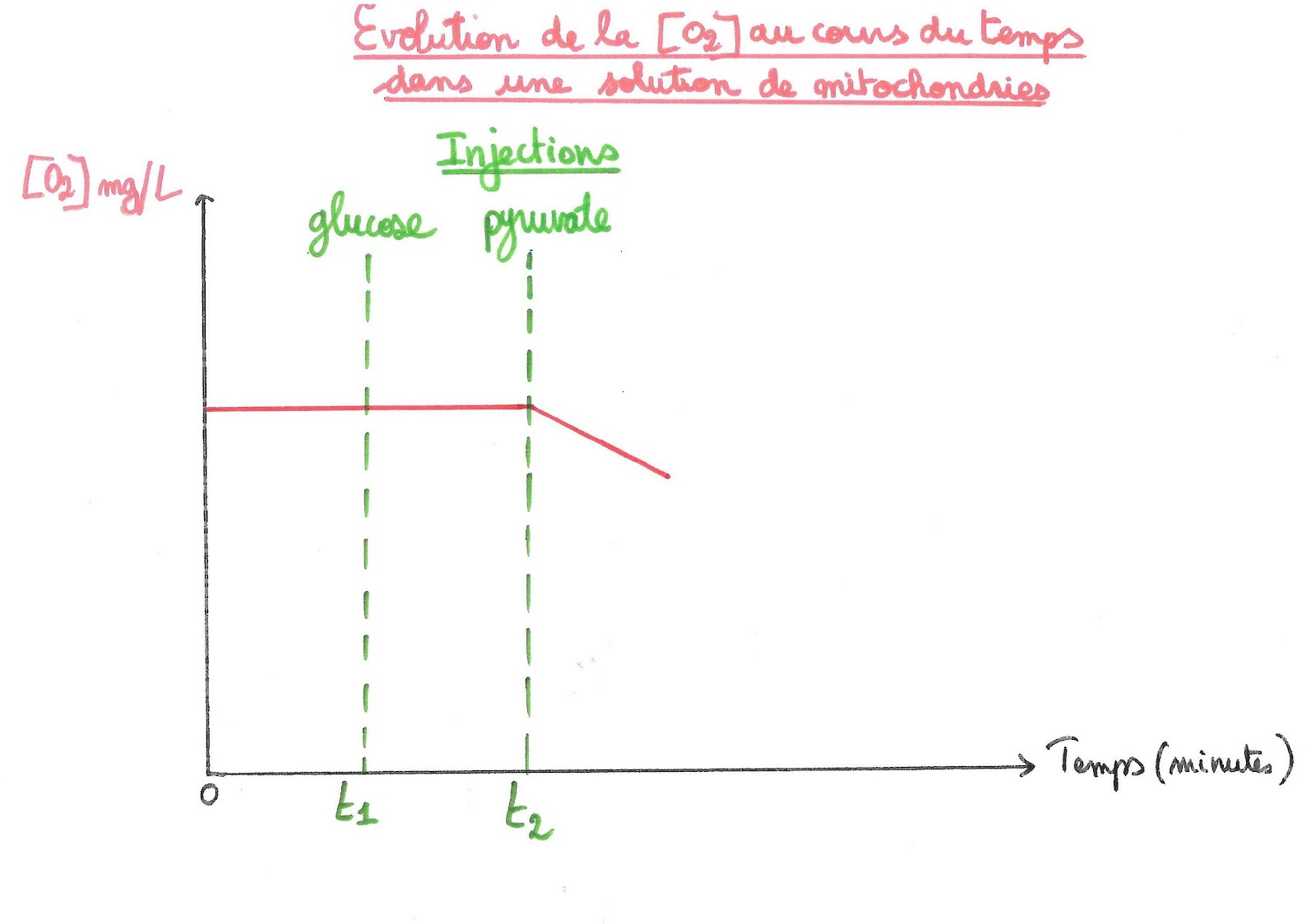
Document 9 : Évolution de la consommation en dioxygène d’une solution de mitochondries en présence de dioxygène après injection de glucose

L’injection de glucose dans une solution de mitochondries ne déclenche pas de consommation de dioxygène. On peut donc dire que la mitochondrie n’utilise pas directement le glucose comme substrat de la respiration. Elle utiliserait donc plutôt le pyruvate, résidu de la glycolyse.
Pour vérifier cela il suffira d’injecter une solution de pyruvate dans la solution pure de mitochondries : s’il y a consommation de dioxygène alors on pourra dire que la mitochondrie utilise le résidu de la glycolyse et donc que cette dernière a bien lieu dans le hyaloplasme de la cellule.
Document 10 : Hypothèse permettant de vérifier l’utilisation du pyruvate par la mitochondrie.

Document 11 : Evolution de la consommation en dioxygène d’une solution de mitochondries en présence de dioxygène après injection de pyruvate
Seule l’injection de pyruvate déclenche une consommation en dioxygène. On peut donc affirmer que la mitochondrie pratique la respiration à partir du résidu de la glycolyse et que cette dernière a entièrement lieu dans le hyaloplasme de la cellule.
2) L’entrée du pyruvate dans un cycle métabolique
Si le glucose sert à régénérer l’ATP, alors le pyruvate le devrait également. Ainsi, si la mitochondrie est bien le lieu de la production d’ATP par respiration, dans la solution de mitochondries pure, en injectant en plus du Pyruvate, de l’ADP et du Phosphate inorganique, on devrait observer une forte consommation de dioxygène au moment de l’injection liée à la production de cet ATP.
Document 12 : Enregistrement de l’évolution de la concentration en dioxygène dans une solution de mitochondries pure après injection de pyruvate et d’ADP + Pi
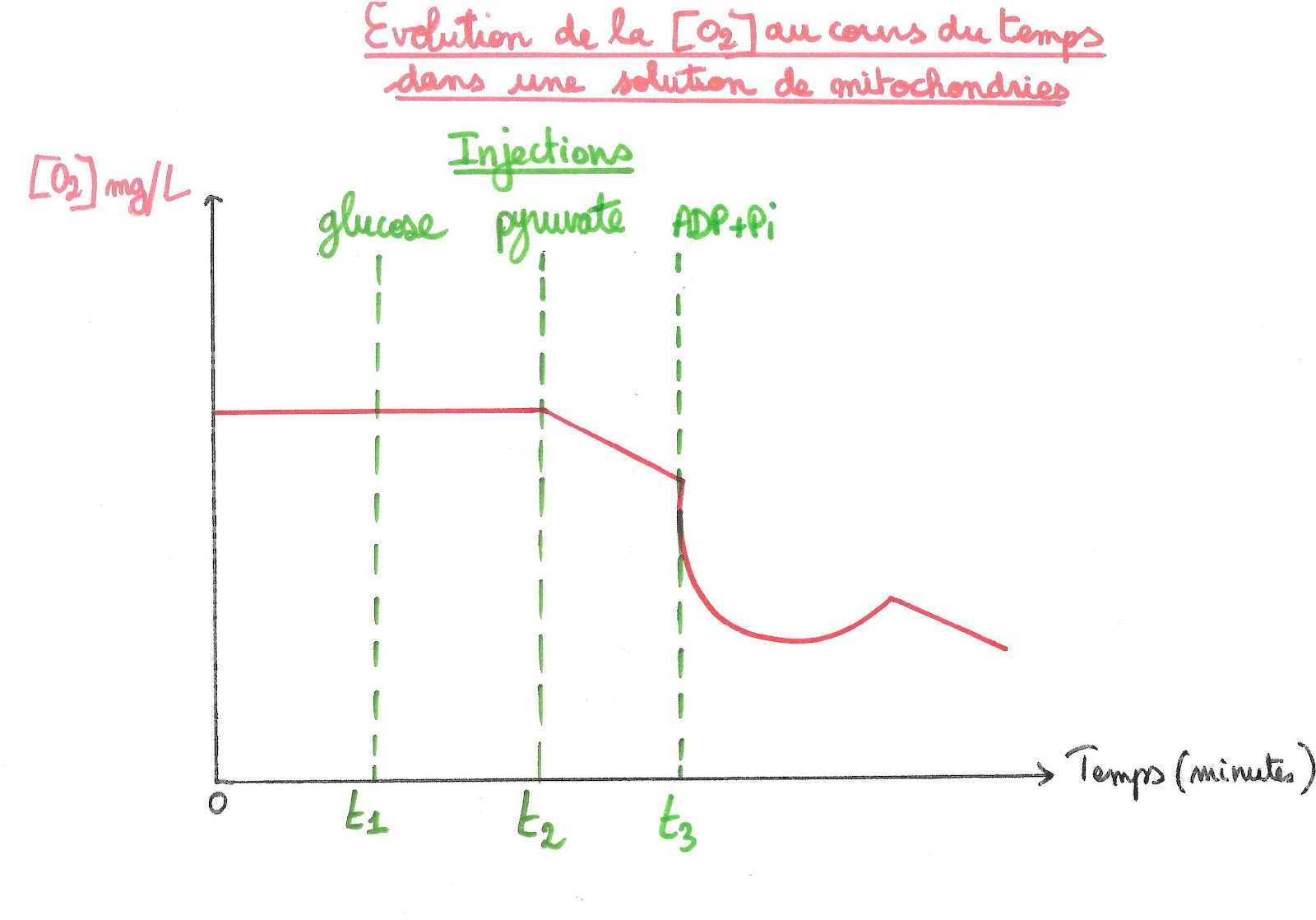
On constate que la consommation en dioxygène augmente fortement quand on injecte en plus du pyruvate, de l’ADP et du Phosphate inorganique. Or on sait que ADP + Pi donne de l’ATP. On en déduit que le pyruvate est utilisé dans des réactions métaboliques qui produisent de l’ATP.
On enregistre dans le même temps, un dégagement de dioxyde de carbone.
Document 13 : Enregistrement de l’évolution de la concentration en dioxygène et en dioxyde de carbone dans une solution de mitochondries pure après injection de pyruvate et d’ADP + Pi

Ainsi dans la matrice des mitochondries, le pyruvate est transformé en CO2. Comme le CO2 ne possède aucun hydrogène, on peut donc penser que les réactions dégradant le pyruvate sont des réactions d’oxydoréduction. On devrait donc trouver des composés réduits RH2 en fin de dégradation.
On utilise le principe de l’absorbance pour déterminer ce qu’il se passe dans la matrice des mitochondries.
Document 14 : Mesures de l’absorbance à 350 nm de différentes solutions et composés
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
On constate qu’en mettant en présence les protéines de la matrice avec le pyruvate et le composé R, on observe une valeur d’absorbance à 250 nm proche de celle des composés réduits RH2.
On en déduit que le R de la solution testée est en train d’être réduit en RH2.
L’oxydation complète du pyruvate en CO2, dans la matrice, est couplée à la réduction du R en RH2.
Ce composé R est également du NAD+. Le pyruvate entre donc dans la mitochondrie et entre dans une série de réactions. Dans un premier temps, il est transformé en une molécule à 2 carbones, l’acétylCoA et en une molécule de CO2. C’est donc une décarboxylation réalisée par le complexe pyruvate déshydrogénase.
Document 15 : Décarboxylation du pyruvate
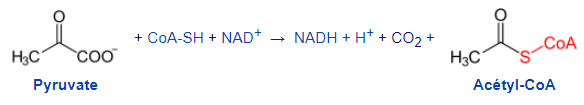
154px-Pyruvat.svg par NEUROtiker prorpe travail, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Pyruvat.svg
176px-Acetyl-CoA-colored.svg, par NEUROtiker prorpe travail, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Acetyl-CoA-colored.svg
Ce dernier va entrer dans un cycle de réactions appelé cycle de Krebs où il va subir une série d’oxydo-réductions.
Document 16 : Cycle de Krebs

Au cours de cette étape, peu d'énergie est encore produite : 1 ATP pour une molécule d’acétylCoA. Par contre 4 composés R sont réduits et on obtient 4 RH2.
Au total, entre la décarboxylation du pyruvate en acétylCoA et le cycle de Krebs, pour 2 pyruvates introduits dans la mitochondrie, il ne se forme que 2 ATP et 10 composés réduits.
L’ensemble de ces réactions n’explique pas la forte consommation de dioxygène ni la forte production d’ATP observées lors de l’injection de pyruvate et d’ADP dans la solution de mitochondries. Il doit donc exister une autre série de réactions qui produira de l’ATP tout en consommant le dioxygène. On peut supposer que cet ensemble de réactions permettra de régénérer le stock de composés R.
C/ La régénération des composés oxydés
Après la glycolyse, la décarboxylation du pyruvate et le cycle de Krebs, la cellule dispose de 6 CO2, 12 RH2 et 4 ATP. Le bilan énergétique n’est pas énorme et il faut recycler les composés réduits.
On peut alors se demander, quel mécanisme est à l’origine de la grande production d’ATP lors de la respiration et comment les composés réduits sont oxydés ?
Les mitochondries n’étant présentes qu’en présence de dioxygène, on peut donc supposer que la production d’ATP et l’oxydation des composés réduits se réalisent au sein de celle-ci.
L’étude au microscope électronique de leur structure révèle la présence, sur les crêtes mitonchondriales, insérées dans la membrane interne, de petites sphères pédonculées, c'est-à-dire des sphères reliées à la membrane par une pièce allongée. Ceci ne s’observe sur aucune autre sorte de membrane.
Pour déterminer le rôle de ces sphères, des morceaux de membrane interne sont obtenus par l’application d’ultrasons sur une mitochondrie. Ces fragments ont tendance à former des vésicules sur lesquelles on va donc observer les sphères pédonculées qui ne seront alors plus en contact avec la matrice mitochondriale mais avec le milieu expérimental. Il est possible de dissocier ces sphères pédonculées des membranes avec de l’urée : les vésicules formées ensuite grâce aux ultrasons n’en porteront donc pas. Le milieu expérimental contient du dioxygène marqué à l’isotope 18, de l’ADP et du Phosphate inorganique Pi. On fait le choix d’y intégrer ou pas des composés réduits RH2.
Document 17 : Première série d’expérimentations sur les sphères pédonculées
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
La comparaison des tests 1 et 2 met en évidence le rôle des sphères pédonculées : elles produisent de l’ATP. En effet, l’ATP est synthétisée en leur présence uniquement. Le test 3 le confirme : quand on rajoute dans le test 2, des sphères pédonculées libres dans le milieu, de l’ATP est produit.
Le test 2 nous permet de plus d'affirmer que c’est la membrane interne de la mitochondrie qui s’occupe de l’oxydation des composés réduits et que le dioxygène est l’accepteur final d’électrons. En effet, en la seule présence de membrane interne mitochondriale, il n’y a pas de synthèse d’ATP mais oxydation des composés réduits et apparition d’une eau lourde.
L’étude du test 4 le confirme et précise la relation entre la membrane et les sphères pédonculées. En effet, en l’absence de membrane (pas de vésicule), il n’y a pas d’oxydation des composés réduits et les sphères pédonculées libres ne produisent pas d’ATP. On peut donc affirmer que la membrane permet l’oxydation des composés réduits et que cette oxydation est la condition nécessaire au fonctionnement des sphères pédonculées. Le test 5 le confirme : en l’absence de composés à oxyder, la sphère pédonculée ne produit pas d’ATP.
On sait que lors de l’oxydation par la membrane interne mitochondriale, des ions hydrogène sont libérés. On peut donc supposer que les protons jouent un rôle dans le fonctionnement des sphères pédonculées. L’oxydation se réalisant dans la membrane interne des mitochondries, on peut donc supposer que des protons s’accumulent dans l’espace inter-membranaire. Or on observe dans le milieu, une formation de molécules d’eau. On peut donc supposer que les protons ont été transférés de l’intérieur des vésicules vers l’extérieur grâce aux sphères pédonculées et que ce transfert est à l’origine de la production d’ATP.
Pour vérifier cela, il suffit de tester le fonctionnement des sphères pédonculées selon le pH intra et extra-vésiculaire et de surveiller ce dernier au cours des tests. En effet, le pH étant le reflet de la teneur en ions hydrogènes, un pH faible traduit une grande concentration en ceux-ci.
Document 18 : Deuxième série d’expérimentations sur les sphères pédonculées
|
|
|
|
|
|
|||
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
La comparaison des tests 1 et 2 permet d’indiquer que les sphères pédonculées produisent de l’ATP uniquement quand le pH intra-vésiculaire est inférieur au pH extra-vésiculaire. La comparaison des tests 2 et 3 permet d’éclaircir le rôle des sphères pédonculées. En effet, c’est seulement dans le test 2, c'est-à-dire en présence de sphères, que les pH intra et extra-vésiculaires se modifient au cours du temps. Le pH diminue dans le milieu extra-vésiculaire et augmente dans le milieu intravésiculaire. On peut donc affirmer que les sphères permettent une sortie des ions H+ de la vésicule et que cette sortie est à l’origine la production d’ATP à partir d’ADP + Pi. Les sphères pédonculées sont donc des pompes à protons synthétisant de l’ATP : on parle d’ATP synthase.
Document 19 : ATP synthase

Atp synthase.PNG par Alex.X via Wikimédia Commons, CC-BY-SA-3.0-migré, modifiée par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Atp_synthase.PNG
Document 20 : Chaîne respiratoire
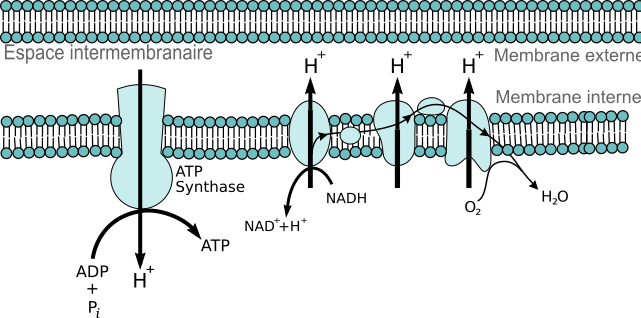
648px-Chaîne_de_transport_des_électrons_mitochondriale.svg par Mitochondrial_electron_transport_chain — Etc4.svg : Fvasconcellos 22:35, 9 septembre 2007 (UTC)travail dérivé: Bob Saint Clar via Wikimédia Commons, domaine publique, modifiée par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Cha%C3%AEne_de_transport_des_%C3%A9lectrons_mitochondriale.svg
En conclusion de ces deux séries de tests, on peut affirmer que la membrane interne mitochondriale possède une chaîne de transporteurs d’électrons à l’origine de l’oxydation des composés réduits et dont l’accepteur final d’électrons est le dioxygène. On parle de chaîne respiratoire. Cette chaîne de transport des électrons est une suite de molécules fixées dans la membrane interne de la mitochondrie. Elles sont disposées dans un ordre décroissant d'affinité pour les électrons. Les électrons provenant des composés réduits se déplacent le long de la membrane sur la chaîne de transport, provoquant au passage l’entrée des protons (H+) libérés, depuis la matrice vers l'espace intermembranaire de la mitochondrie, créant ainsi un gradient (une différence de concentration). L'accumulation de protons fait fonctionner l'ATP synthase (une pompe à protons) qui va profiter du retour de 3 de ces protons dans la matrice par son canal central pour associer de l'ADP et une molécule de phosphate inorganique, et fabriquer ainsi une molécule d'ATP. L'accepteur final d'électrons de cette chaîne de transport étant le dioxygène, ce dernier, en se liant à deux protons, forme par réduction, une molécule d'eau dans la matrice mitochondriale. L’oxydation des 10 composés réduits RH2 en composé oxydé R par la chaîne respiratoire produit 32 ATP.
La production d’ATP de la voie aérobie s’élève à 36 ATP par molécule de glucose : 2 ATP provenant de la glycolyse, 2ATP du cycle de Krebs et 32 ATP de la chaîne respiratoire.
Document 21 : Schéma bilan de la respiration cellulaire

{kind=link}
{kind=link}
Glycolyse et voie aérobie - SVT - SANTÉ Term spé #9 - Mathrix
Date de dernière mise à jour : 07/05/2024