Les lois de Mendel
vidéo en bas de page ^^
I Les lois de Mendel
Document 1 : Gregor Mendel

Source : Gregor-mendel-39282-1-402.jpg par Sanjana Kadur via Wikimédia Commons, Creative Commons Attribution-Share Alike 4.0 International, https://commons.m.wikimedia.org/wiki/File:Gregor-mendel-39282-1-402.jpg
Gregor Mendel né le 20 juillet 1822 et mort le 6 janvier 1884, était un moine catholique du monastère Saint-Thomas de Brünn située en Tchéquie. Botaniste et généticien, il est communément reconnu comme le père fondateur de la génétique. Il est à l’origine de ce qui est actuellement appelé les lois de Mendel et qui définissent la manière dont les gènes se transmettent de génération en génération.
À son époque, comme un enfant présentait à la fois des caractères de son père et de sa mère, il était communément admis que l’hérédité se faisait par mélange des caractères parentaux dans certaines proportions. Mendel réfuta cette loi de l’hérédité par mélange.
Document 2 : Principe de l’hérédité par mélange

Source : Blending Inheritance fr.svg par Raminagrobis via Wikimédia Commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Blending_Inheritance_fr.svg
Il travailla sur plusieurs caractères de lignées de petits pois et émit 3 lois.
- la loi d’uniformité des hybrides de première génération
- la loi de ségrégation des caractères dans la génération F2 (appelée aujourd’hui loi de disjonction des allèles)
- la loi d’indépendance de la transmission des caractères ;
Le choix du Pois fût très judicieux car il existe pour cette plante dont le temps de régénération est court et la descendance nombreuse, un grand nombre de couples de caractères variants bien définis (couleur et forme des fruits et des graines mûres, couleur des pétales, taille de la plante, etc).
Document 3 : Caractères du pois étudiés par Mendel
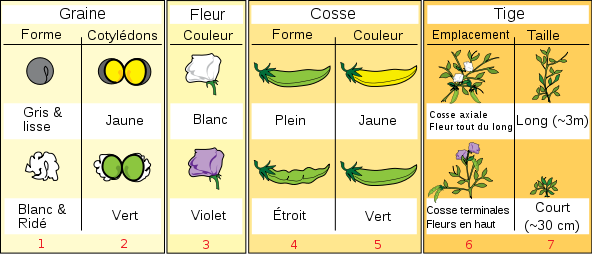
Source : Mendel sept caractères fr.svg, par Mariana Ruiz LadyofHats (traduction française Moez ) via Wikimédia Commons, Domaine publique https://commons.wikimedia.org/wiki/File:Mendel_seven_characters_fr.svg
La structure de la fleur permet l'expérimentation :
- les étamines et les pistils étant enfermés dans la carène de la fleur (gaine formée par 2 pétales soudés), l'autofécondation de la plante est possible.
- on peut enlever les étamines avant que le pollen ne soit mûr et féconder le pistil avec du pollen prélevé sur une autre plante (fécondation croisée).
Document 4 : Fleur de petit pois dévoilant la carène en son centre

©RS.2019
Mendel a travaillé sur des plants de petits pois présentant soit des graines lisses soit des graines ridées. Les individus utilisés étaient des individus de souches pures car isus d'une lignée présentant ce caractère depuis plusieurs générations. Il constata systématiquement qu’en génération hybride F1, c’est-à-dire la génération issue de la fécondation n°1 des gamètes de parents P1 (plant à graines lisses) et P2 (plant à graines ridées), aucun individu ne présentait de mélange de caractères. Tous les individus F1 présentaient une uniformité des caractères correspondants aux caractères d’un seul des deux parents (P1) : ils ne présentaient que l’aspect lisse des graines. Le caractère « pois lisse » était donc dominant sur le caractère « pois ridé ». C’est la « loi d’uniformité des hybrides de première génération ».
Document 5 : Croisement d’un petit pois à graines lisses avec un petit pois à graines ridées
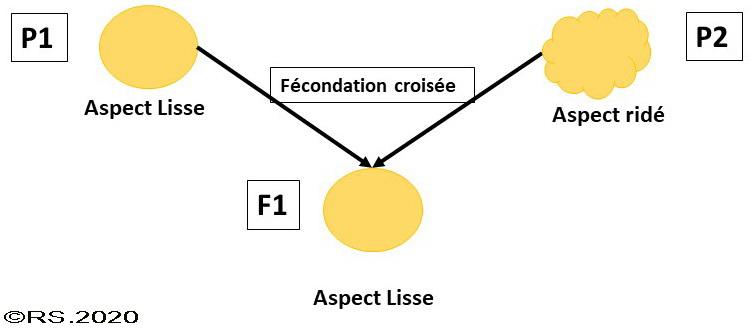
Quel que soit le caractère étudié (couleur de la graine ou des fleurs, aspect de la graine….) à chaque fois un caractère semblait sauter une génération.
En effet, en constatant que dans la génération F2 issue du croisement de deux individus F1, apparaissait un caractère absent en F1 mais présent chez P2, il mit en évidence que l’hérédité était transmise par une « particule », un élément porteur d’information appelé aujourd’hui « allèle ». Chaque plant étant issu de deux parents, il possède forcément les particules en double exemplaire. Les deux particules se séparent (ségrégent) lors de la formation des gamètes de sorte que chaque gamète ne possède qu’une particule sur les 2. C’est la « loi de ségrégation des caractères dans la génération F2 » appelée aujourd’hui loi de disjonction des allèles.
Document 6 : Croisement de petits pois de lignée pure P1 et P2

Pour Mendel, dans le cas de l’étude de 2 caractères, les plants de petits pois à graines jaunes et lisses étant de souches pures, ils possèdent un couple de particules dominantes codées « A » pour la couleur jaune et un couple de particules dominantes codées « B » pour le caractère lisse alors que les plants de petits pois à graines vertes et ridées possèdent un couple de particules récessives (non dominantes) codées « a » pour la couleur verte et un couple de particules récessive codées « b » pour le caractère ridé. Ainsi au moment de la formation des gamètes, chacun de ces derniers ne contient qu’une seule particule d’un couple. Leur croisement donne toujours une génération F1 où tous les individus ont des pois jaunes (particules A et a) à graines lisses (particules B et b).
Document 7 : Couples de particules portées par les petits pois jaunes et lisses et par les petits pois verts et ridés.

En travaillant sur 2 caractères à la fois (couleur et aspect de la graine), il a constaté que les particules associées aux deux caractères n’étaient pas forcément transmises ensemble des parents aux descendants et qu’elles pouvaient se disjoindre pour s’associer, faisant apparaître de nouvelles combinaisons de caractères ou phénotypes en F2. C’est « la loi d’indépendance de la transmission des caractères » : la ségrégation d'un couple d'allèles est indépendante de celle d'un autre couple d'allèles.
Document 8 : Résultat du croisement de petits pois F1 (AaBb) entre eux

II Le rôle de la fécondation
La fécondation est l’union de deux gamètes dits haploïdes car ils ne contiennent qu’un seul exemplaire de chaque sorte de chromosomes. La réunification des deux noyaux appelés caryogamie, rétablit le caryotype et donc rétablit la diploïdie. Ainsi le cycle de vie de l’Homme par exemple est une alternance de phases diploïdes et haploïdes où la phase diploïde est prédominante. La fécondation permet ainsi la stabilité du caryotype au fur et à mesure des générations.
Document 9 : Cycle de vie de l’Homme

©RS.2019
Si le caryotype est stable, il n’en reste pas moins que les individus restent uniques du fait que chaque gène existe sous différentes versions appelées « allèles ».
Ainsi pour un gène donné, un individu peut être qualifié d’homozygote s’il possède deux allèles identiques pour ce gène. Si cet allèle est un allèle dominant on parlera d’individu homozygote dominant. Si l’allèle est récessif on parlera d’individu homozygote récessif. Un individu recevant deux allèles différents sera qualifié d’hétérozygote.
Document 10 : Position des gènes et des allèles chez des individus différents.
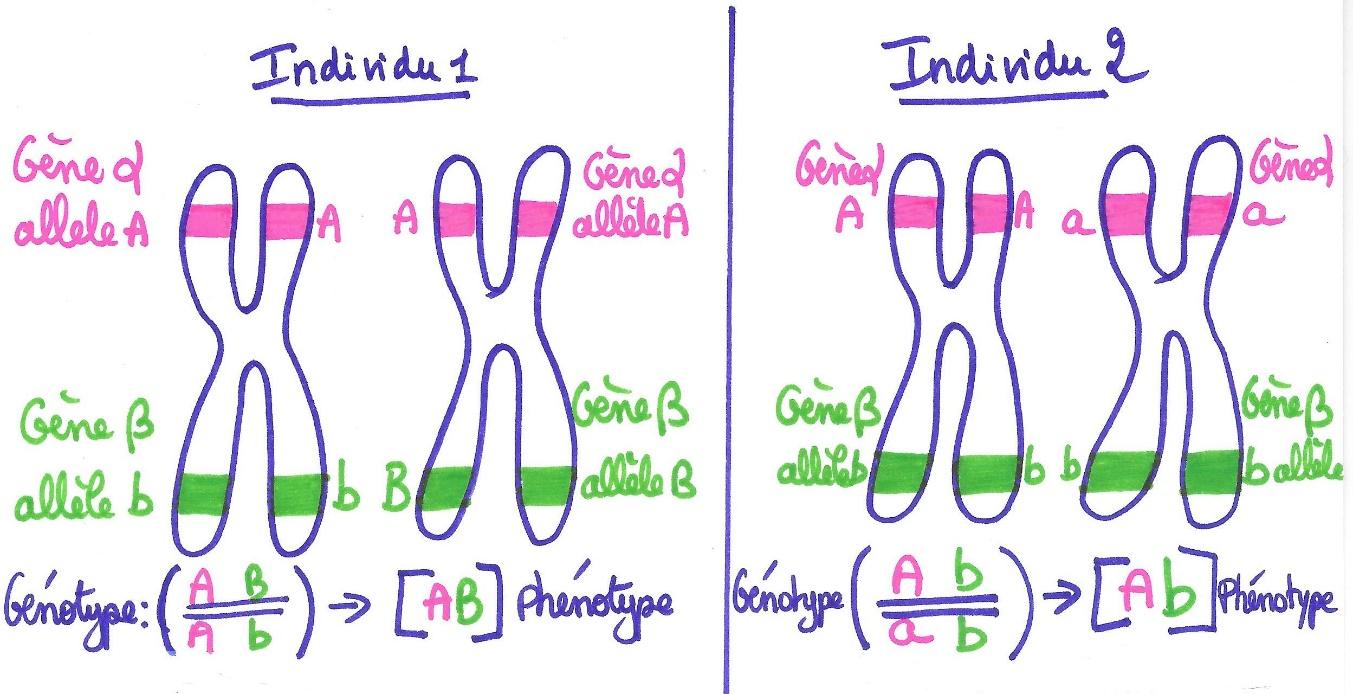
Les gènes étudiés sur le schéma précédent, les gènes alpha et bêta, sont des gènes situés sur le même chromosome : on dit qu’ils sont liés. L’individu 1 est homozygote dominant pour le gène alpha et hétérozygote pour le gène bêta. L’individu 2 est hétérozygote pour le gène alpha et homozygote récessif pour le gène bêta.
Précisions : Dans le système de codification en génétique, le génotype (ensemble des gènes que l’on possède) s’écrit toujours entre parenthèses alors que le phénotype (ensemble des caractères que l’on présente) s’écrit entre crochets. La lettre majuscule représente l’allèle dominant et la lettre minuscule l’allèle récessif du gène d’intérêt. Les doubles slashs représentent la paire de chromosomes homologues. Quand deux gènes sont portés par des chromosomes différents (on dit qu’ils sont indépendants), il convient de placer une virgule entre les codes utilisés pour les gènes. Quand les gènes sont portés par le même chromosome (on dit qu’ils sont liés), on ne met pas de virgule et les lettres utilisées pour coder les gènes sont écrites l’une à côté de l’autre. Si l’on doit écrire le génotype d’un gamète, on n’écrira donc entre parenthèses qu’une seule lettre associée à un seul slash (A/) car le gamète ne possède qu’un seul exemplaire de chaque sorte de chromosomes. Les gamètes de l’individu 1 seront donc de type (Ab/) ou (AB/) et ceux de l’individu 2 seront (Ab/) ou (aB/).
Dans un croisement, il est possible d’anticiper les résultats de celui-ci en établissant un tableau de croisement appelé également tableau de fécondation, à condition de connaître précisément le génotype des parents.
Considérons deux individus de phénotype [AB]. Les gènes responsables des caractères sont situés sur des chromosomes différents. Considérons ces individus comme hétérozygotes : leur génotype est donc (A//a, B//b).
Ainsi il existe pour chaque individu quatre sortes de gamètes différents : (A/,B/) , (A/, b/) , (a/, B/) , (a/,b/).
Document 11 : Différents gamètes possibles pour un individu hétérozygote pour 2 gènes indépendants.
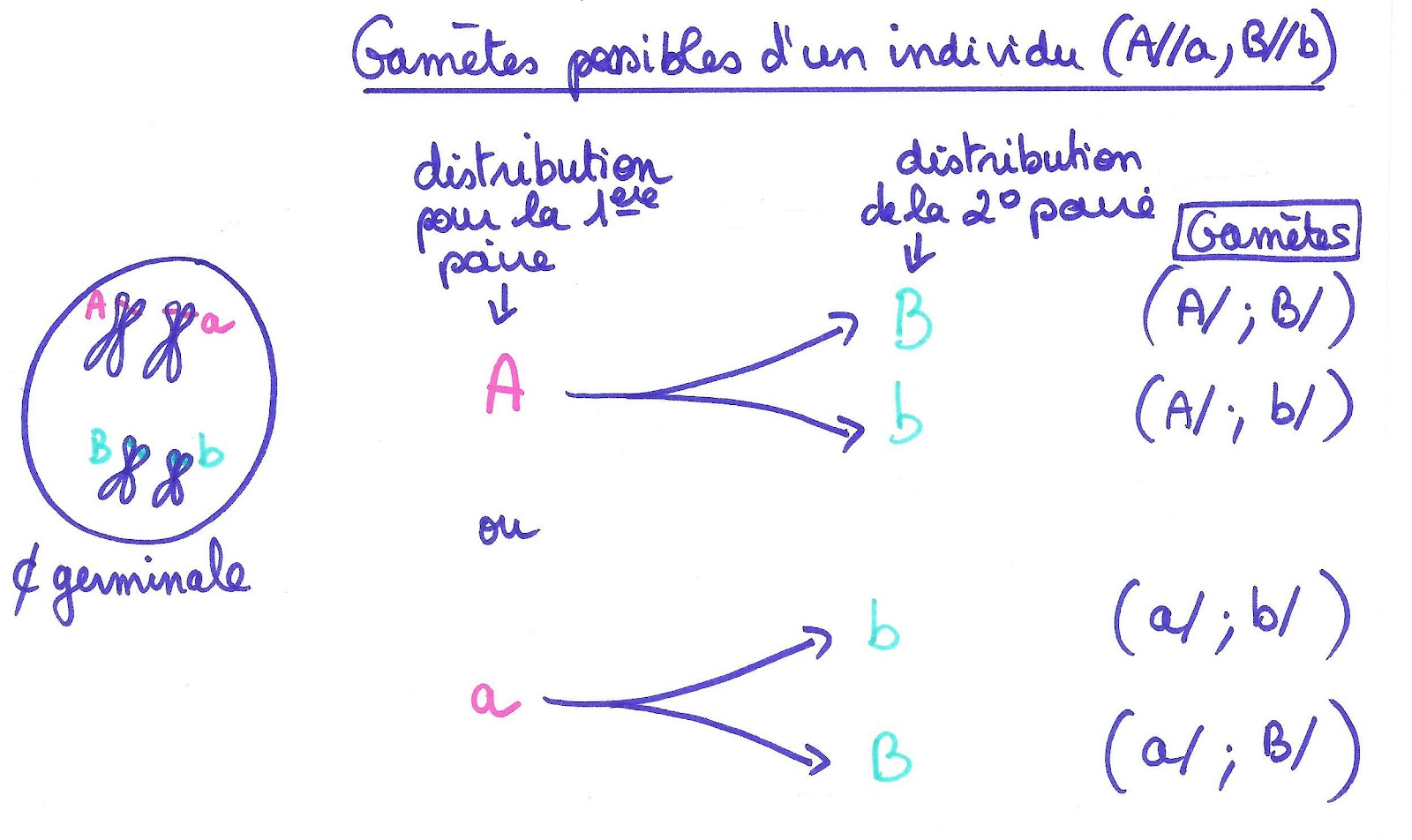
Le tableau de croisement pour ces deux individus hétérozygotes pour 2 gènes indépendants permettra de réunir les différentes sortes de gamètes entre eux et de déterminer ainsi tous les génotypes possibles et donc les phénotypes possibles de leurs descendants ainsi que leurs proportions respectives.
Document 12 : Différents gamètes possibles pour un individu hétérozygote pour 2 gènes indépendants.
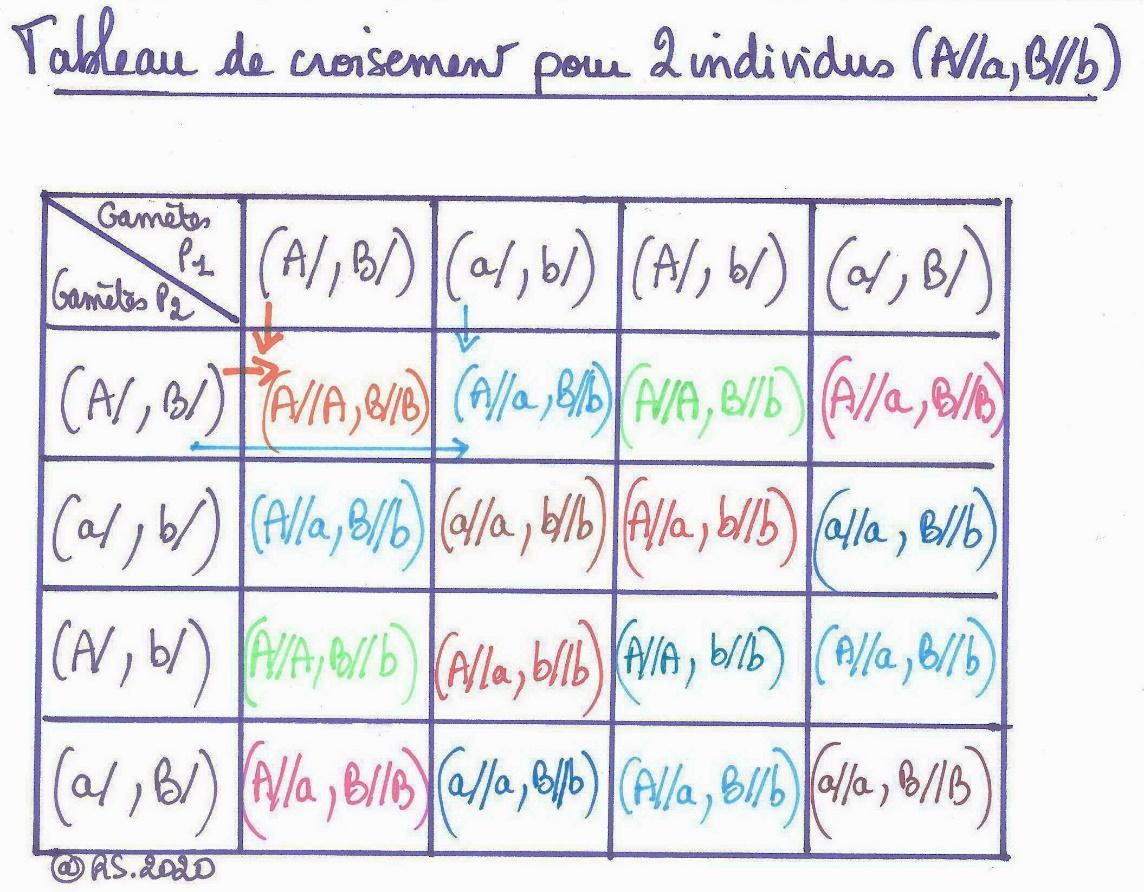
On remarque ainsi, qu’en considérant 2 gènes, il y a 4 possibilités de gamètes différents à l’origine de 9 génotypes de fœtus différents. La fécondation est donc source de diversité.
III utilisation des lois de Mendel pour définir le génotype d’un individu
A/ Produire un phénotype recherché
Mendel avait mis en évidence l’existence d’une dominance et d’une récessivité des allèles en étudiant des caractères des graines de petits pois mais également en étudiant les couleurs des fleurs de petits pois. En effet, en croisant un plan aux fleurs roses (P1) avec un plan aux fleurs blanches (P2), la génération F1 présentait systématiquement des fleurs roses. Ainsi deux allèles ont été mis en évidence : l’allèle B responsable du caractère rose qui est dominant sur l’allèle b responsable du caractère blanc qui lui est donc récessif (il ne s’exprimera que s’il est présent en deux exemplaires). C’est la 1ère loi de Mendel, la loi d’uniformité des hybrides de première génération.
Document 13 : Étude de la transmission du caractère « couleur de la fleur » chez le petit pois
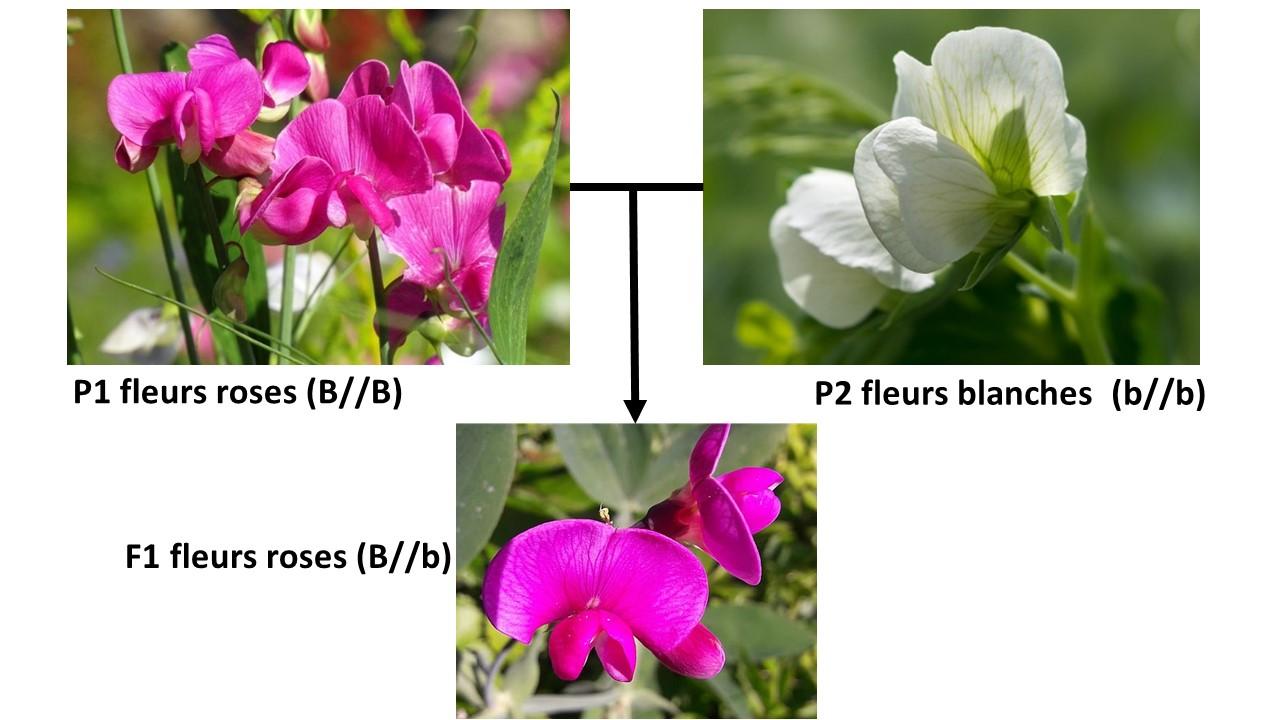
Sources :
P1 : fleur de pois rose sucré, par Nowaja via Pixabay, Pixabay licence, https://pixabay.com/fr/photos/pois-sucr%C3%A9-parfum%C3%A9-rose-4551745/
P2 : fleur de pois blanche : Didgeman, via Pixabay, Pixabay Licence, https://pixabay.com/fr/photos/pois-fleur-de-pois-blanc-fleur-5215863/
F1 : sweet-pea-sweet-peas-pink-beautiful-lathyrus-odoratus-flowers-summer-color-colors,CC0, https://www.pikist.com/free-photo-velzx/fr
Cependant si Mendel avait étudié d’autres plantes, il aurait pu constater que parfois en génération F1 apparaissent des fleurs dont la couleur n’existe ni chez le parent 1 ni chez le parent 2 et semble provenir d’un mélange des deux. C’est le cas par exemple chez les roses où le croisement d’une rose rouge et d’une rose blanche donne une couleur rose. Ce caractère traduit une dominance incomplète de chacun des deux allèles : ils s’expriment tous les deux, on parle de codominance.
Document 14 : Étude de la transmission du caractère « couleur de la fleur » chez la rose

Sources :
P1 : rose rouge, par GoranH, via Pixabay, Pixabay licence, https://pixabay.com/fr/photos/artificielle-rose-rouge-fleur-4933142/
P2 : rose blanche par Jolenka, via Pixabay, Pixabay licence, https://pixabay.com/fr/photos/la-rose-blanche-toussaint-2907862/
F1 : rose rose clair par PurpleOwl, via Pixabay, Pixabay licence, https://pixabay.com/fr/photos/rose-rose-rose-rose-clair-fleur-5227900/
Aujourd’hui on utilise les connaissances en génétiques afin d’orienter les productions de plantes à caractères spécifiques correspondant à la demande des consommateurs. Le même travail est effectué dans les élevages afin de produire des animaux aux caractéristiques utiles à l’Homme et voulues par lui.
B/ Prévoir les différents phénotypes possibles
En utilisant la 2ème loi de Mendel, la ségrégation des caractères en génération F2, c'est-à-dire en considérant le fait que les allèles se disjoignent au moment de la fabrication des gamètes, il est possible d’envisager la descendance possible d’un couple en effectuant un tableau de croisement.
On peut citer comme exemple la prévision du groupe sanguin d’un fœtus chez un couple de parents. Il existe 4 groupes sanguins.
Document 15 : Le caractère « groupe sanguin » chez l’Homme
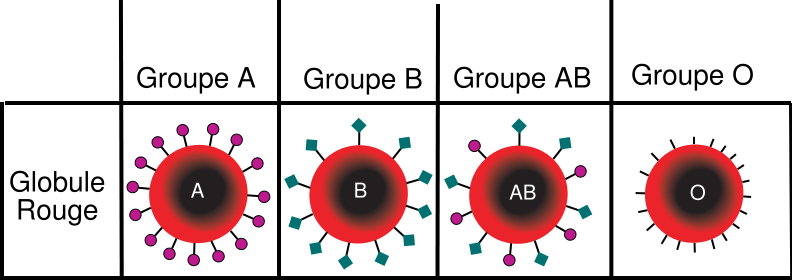
Source : Groupes sanguins.png par Amouna2000 via Wikimédia Commons, domaine publique, modifié par Sandra rivièrehttps://commons.wikimedia.org/wiki/File:Groupe_sanguin_ABO.svg
Le gène responsable du groupe sanguin existe trois versions différentes : l’allèle A, l’allèle B et l’allèle O.
L’allèle A est responsable de la production d’une molécule de type A en surface des globules rouges. L’allèle B est responsable de la production d’une molécule de type B alors que l’allèle O n’induit la fabrication d’aucune molécule à la surface des globules rouges. L’allèle O est récessif ce qui signifie qu’il ne s’exprime que s’il est « seul » c’est-à-dire non face à l’allèle A ou B et donc uniquement s’il est présent en double exemplaire. Les allèles A et B sont donc dominant sur l’allèle O. Cependant un individu de génotype (A//B) présentera les deux types de molécules à la surface de ses globules rouges : il sera donc du groupe [AB]. Nous sommes dans le cas d’une dominance incomplète appelée également codominance. Ainsi un individu de phénotype [A] peut présenter deux génotypes : (A//A) ou (A//O). Il en va de même pour le phénotype [B].
Document 16 : Phénotypes sanguins et génotypes associés
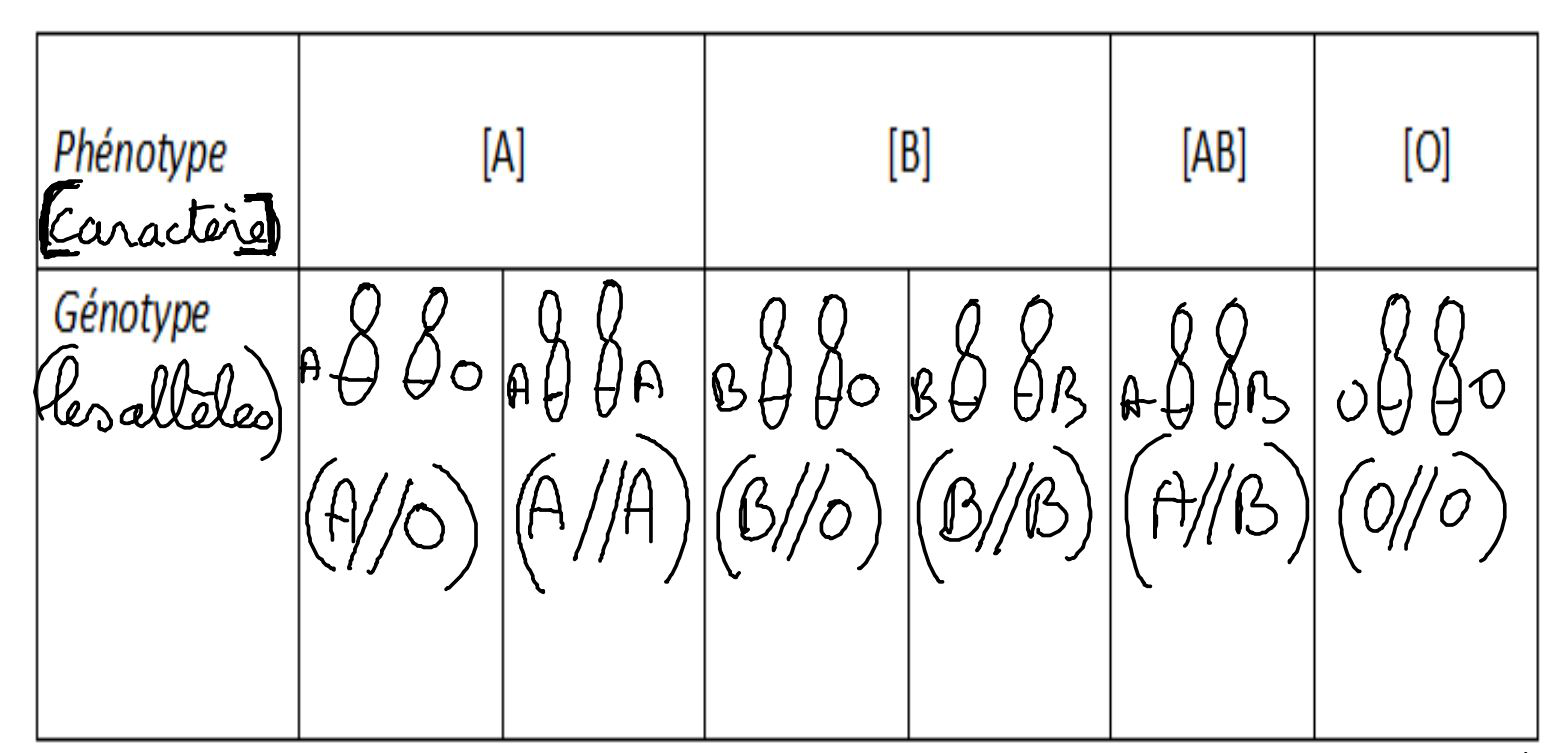
La connaissance du phénotype des parents à elle seule ne permet pas de réaliser la détermination des génotypes possibles d’un individu à venir. La connaissance du génotype de ceux-ci est importante. Elle permet la réalisation d’un tableau de croisement afin d’estimer la probabilité pour un descendant F1 de présenter un phénotype d’intérêt.
Ainsi par exemple dans le cas de parents hétérozygotes de groupe [A] et [B], des individus F1 auront quatre possibilités différentes de phénotypes : [AB], [A] ; [B] et [O] alors que dans le cas de parents homozygotes de groupe [A] et [B], des individus de la génération F1 ne pourront présenter qu’un seul phénotype, le phénotype [AB].
Document 17 : Tableau de croisement pour des parents hétérozygotes de groupe [A] et [B]

©RS.2020
Document 18 : Tableau de croisement pour des parents homozygotes de groupe [A] et [B]

{kind=link}
©RS.2020
Lois de Mendel - SVT - LA VIE Term spé # 4- Mathrix
Date de dernière mise à jour : 22/05/2021