Le devenir des produits de la photosynthèse
Vidéo en bas de page ^^
I Les molécules qui contribuent à la croissance et au port de la plante
Contrairement aux animaux qui possèdent un appareil squelettique pour certains ou un exosquelette rigide pour d’autres, les végétaux ne possèdent pas d’organes spécialisés dans le port de leur organisme. Ce sont les parois cellulaires qui à l’instar d’une brique en béton vont permettre aux tissus d’être plus ou moins rigides et de maintenir le port des plantes.
La paroi des cellules végétales est principalement constituée de cellulose et peut parfois être renforcée par d’autres molécules complexes appelées lignines.
A/ La cellulose
La cellulose constitue 35 à 50 % de la biomasse végétale formant ainsi la première famille des composés par ordre d’abondance dans les plantes et dans les écosystèmes terrestres où domine la biomasse végétale morte ou vive.
La cellulose est un glucide complexe constitué de chaînes linéaires de molécules de glucose. Sa formule est donc (C6H10O5)n . Le motif répété est donc le C6H10O5 et il est répété un très grand nombre de fois, voilà pourquoi on rajoute des parenthèses et on indique « n » en indice.
C’est une enzyme appelée cellulose synthase qui va permettre cette polymérisation et qui n’existe que chez les bactéries et les plantes. Dans un premier temps deux unités glucose s’associent pour former une molécule appelée cellobiose, puis les cellobioses s’associent entre elles pour former la macromolécule de cellulose.
Les macromolécules de cellulose s’associent entre elles pour former des microfibrilles qui elles-mêmes s’associent pour former des feuillets rigides et résistants. Ces feuillets se superposent et forment les parois des cellules végétales. Il s’établit des liaisons hydrogènes entre les molécules de glucose des différentes chaînes, ce qui permet une résistance importante des parois.
Document 1 : Association de chaînes de cellulose par des liaisons hydrogènes.

Source : Liaisons hydrogène entre molécules de cellulose.svg, par Laghi.l, via wikimédia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Liaisons_hydrog%C3%A8ne_entre_mol%C3%A9cules_de_cellulose.svg
La paroi des cellules végétales est complexe. Elle est formée par l’association de microfibrilles de cellulose accolées d’un côté à la membrane de la cellule et de l’autre à la lame moyenne, structure riche en pectine commune aux deux cellules voisines.
Document 2 : Structure de la paroi des cellules végétales.

Source : Diagramme de paroi cellulaire végétale-en.svg Par LadyofHats, Via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Plant_cell_wall_diagram-en.svg
La cellulose étant le principal constituant des parois des cellules, elle est donc synthétisée en abondance pendant la division cellulaire et de fait lors de la croissance du végétale. Elle constitue une protection pour la cellule et la rend plus résistante.
B/ La lignine
La lignine n’est pas à proprement parler une molécule. C’est un réseau tridimensionnel hydrophobe complexe constitué de composés phytochimiques de la famille des monolignols. Il n’y a pas une lignine mais des lignines. En effet les lignines sont des polymères de monolignols dont on trouve trois catégories dans des proportions plus ou moins importantes : sont l’alcool coumarylique l’alcool coniférylique, et l’alcool sinapylique. L’alcool coniférylique est très abondant (80 %) chez les conifères et l’alcool sinapylique (50 %) l’est chez les angiospermes.
Document 3 : Molécules de monolignols à l’origine des molécules de lignine.
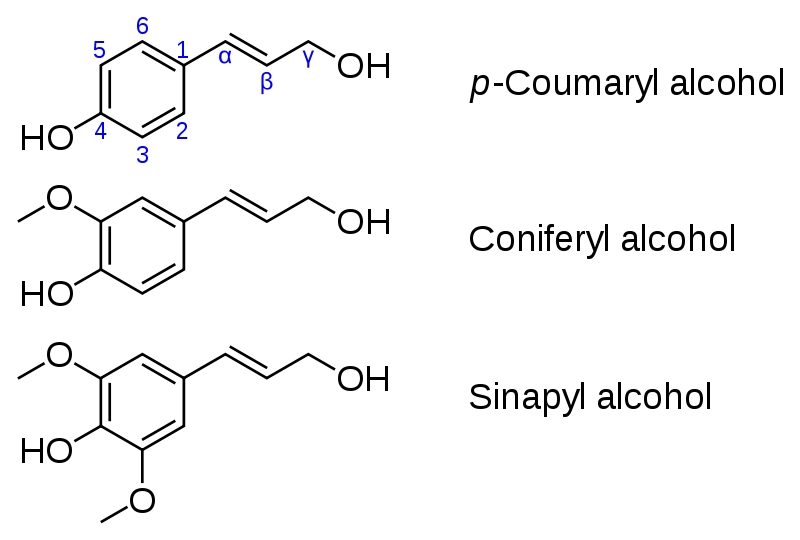
Source : Monolignols.svg, par
Fvasconcellos 21:51, 4 janvier 2008 (UTC). Image: MonolignolStructure.jpg par Chino .via Wikimedia commons, domaine publique, https://commons.wikimedia.org/wiki/File:Monolignols.svg
Document 4 : Diagramme des voies de sécrétion, y compris le noyau, le réticulum endoplasmique et l'appareil de Golgi.
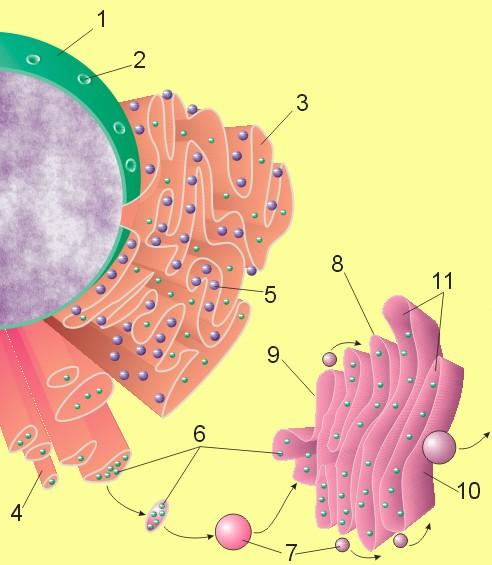
Légende : Membrane nucléaire
- Pore ??nucléaire
- Réticulum endoplasmique rugueux (rER)
- Réticulum endoplasmique lisse (sER)
- Ribosome attaché au rER
- Macromolécules
- Vésicules de transport
- Appareil de Golgi
- Face Cis de l'appareil de Golgi
- Face trans de l'appareil de Golgi
- Citernes de l'appareil de Golgi
Source : Nucleus ER golgi.jpg par Magnus Manske via Wikimedia commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Nucleus_ER_golgi.jpg
Ces monolignols sont obtenus notamment par la biotransformation d’un acide aminé produit lors de la photosynthèse, la phénylalanine, grâce à une enzyme, la phénylalanine ammonia-lyase (PAL). Les monolignols sont synthétisés dans un organite particulier du cytoplasme, le réticulum endoplasmique. Ils sont enfermés dans des vésicules dirigées vers l’appareil de golgi pour y être modifiés : du glucose leur est ajouté pour les rendre hydrosolubles et pour réduire leur toxicité. Ils sont alors exocytés à travers la paroi cellulaire vers le contenu extracellulaire appelée apoplasme. Le glucose est alors retiré et les monolignols sont polymérisés en lignine dans la paroi de la cellule végétale. La lignine est donc déposée secondairement dans la paroi de certaines cellules, c’est-à-dire bien souvent après leur spécialisation.
La lignine possède un pouvoir d’imperméabilisation du fait de son hydrophobicité. Ainsi on la trouve dans les cellules de tissus servant au transport de l’eau et des sels minéraux, en l’occurrence dans les parois des cellules constitutives du xylème.
La lignine confère une grande rigidité aux cellules végétales qui en possèdent. De manière générale, les plantes herbacées ont un port souple indiquant qu’elles possèdent globalement peu de lignine contrairement aux arbres dont le tronc en est très riche.
Document 5 : Coupe transversale de tige de petit pois (plante type herbacée) MO x400
Les tissus ligneux correspondent au xylème éparpillé en petits faisceaux dans la tige. La plante herbacée est souple et ne résiste pas à l’hiver.

L’étude d’une coupe transversale de tige de tilleul met en évidence un grand nombre de cellules dont les parois sont riches en lignines.
Document 6 : Coupe transversale d'une tige de tilleul, colorée au carmin MOx400

Source : Cork cambium 1.jpg par Kje4532 — Travail personnel, via Wikimédia Commons, CC BY-SA 4.0, modifié par Sandra Rivière https://commons.wikimedia.org/wiki/File:Cork_cambium_1.jpg
L’existence de mutants de la voie de biosynthèse des monolignols chez Arabidospsis thaliana prouvent le rôle de la lignine dans le port des végétaux. En effet, plusieurs gènes mutés sont responsables d’une diminution de la lignification des parois du xylème des plantes provoquant des collapsus (effondrement des vaisseaux) et de fait, un port retombant de la plante
II Les molécules de réserves
Au printemps et en été, la production de matière organique par les végétaux chlorophylliens est maximale. La photosynthèse est réalisée principalement dans le parenchyme chlorophyllien des feuilles. Le sucre produit sert à la fois de source de matière et d’énergie pour la production de matière organique. La matière produite va être distribuée aux parties non chlorophylliennes et donc hétérotrophes de la plante : racines, fleurs, fruits....
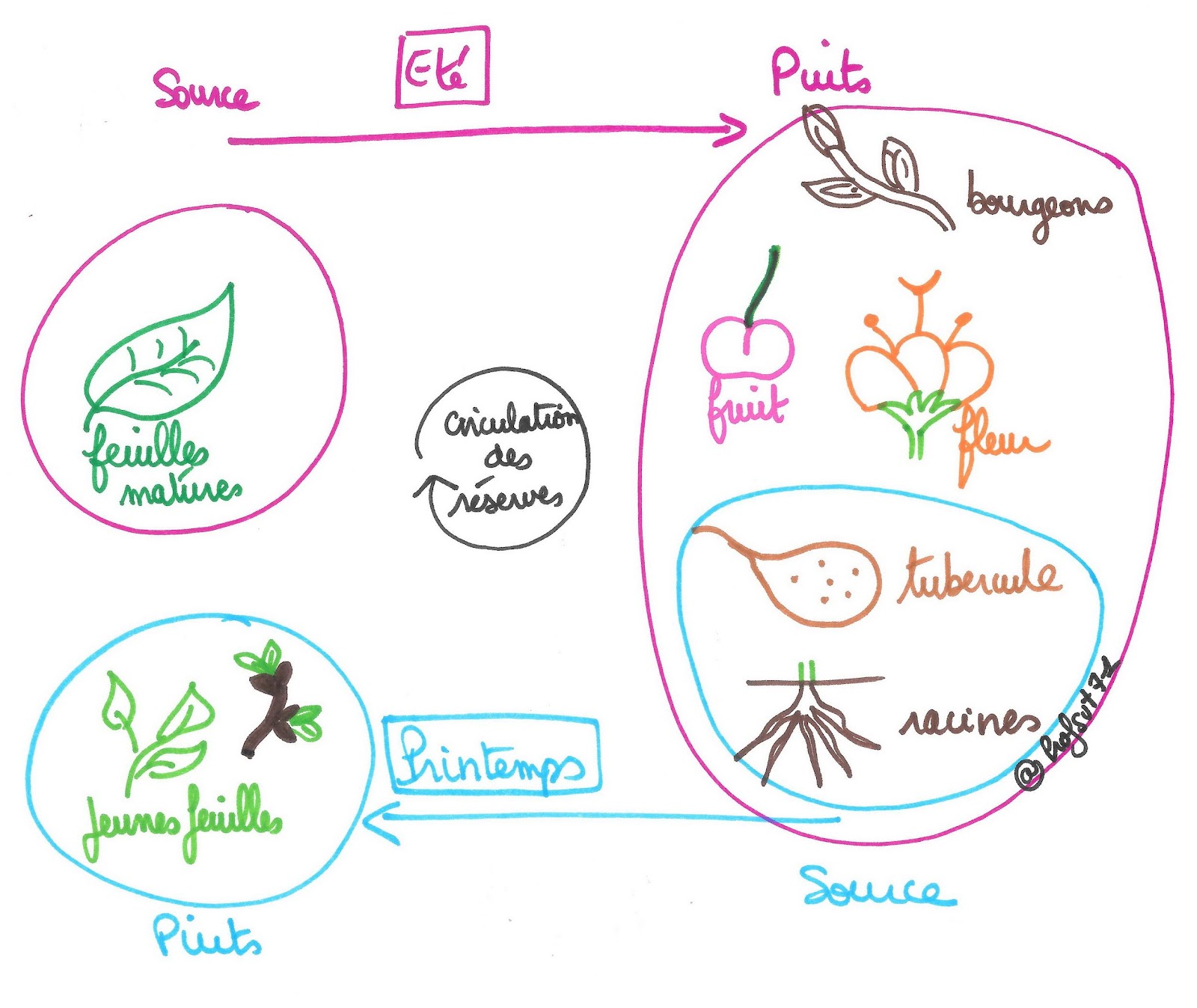
Concernant la production et la consommation ou le stockage de matières on parle d’organe-source et d’organe-puits.
Un organe-source est un organe producteur de matière : ce sont les feuilles en été.
Un organe-puits est un organe consommateur de matière dans le but de l’utiliser ou de la stocker : ce sont les bourgeons, les fleurs, les fruits, les racines et les organes de réserve type tubercules en été. La circulation de la matière organique au sein du phloème se fait par différence de concentration des substances entre les organes-sources et les organes-puits.
Document 7 : Mécanisme de circulation de matière entre organes-sources et organes-puits

Les organes-sources et les organes-puits peuvent changer selon les saisons. Ainsi au début du printemps, les organes-sources ne sont plus les feuilles mais certains organes de réserve, et les organes-puits sont alors les bourgeons et les jeunes feuilles.
Document 8 : Circulation de la matière entre organes-sources et organes-puits.
On va distinguer deux catégories de réserves :
- les réserves journalières localisées dans les vacuoles, les chloroplastes et le cytoplasme des cellules
- les réserves saisonnières dans des tissus et organes spécialisés des végétaux qui accumulent de la matière pendant la belle saison.
Les organes de réserve des végétaux ont 2 fonctions importantes :
- la reprise de la vie active après l’hiver ou une saison particulièrement sèche
- l’optimisation de la reproduction, qu’elle soit asexuée ou sexuée et dans ce dernier cas, les réserves concernent soit le futur individu c’est-à-dire la graine, soit l’organe de dissémination de celui-ci.
Concernant le passage de la mauvaise saison on peut citer en exemple les plantes bisannuelles comme la carotte ou l’oignon. La carotte est une racine qui va se gorger de sucre le premier été pour résister à l’hiver et produire ses fleurs au printemps suivant. L’oignon est également une plante bisannuelle cependant ses réserves également sucrées vont se localiser dans des feuilles charnues constituant un bulbe.
Document 9 : Cycle bisannuel de la carotte.

©RS.2020
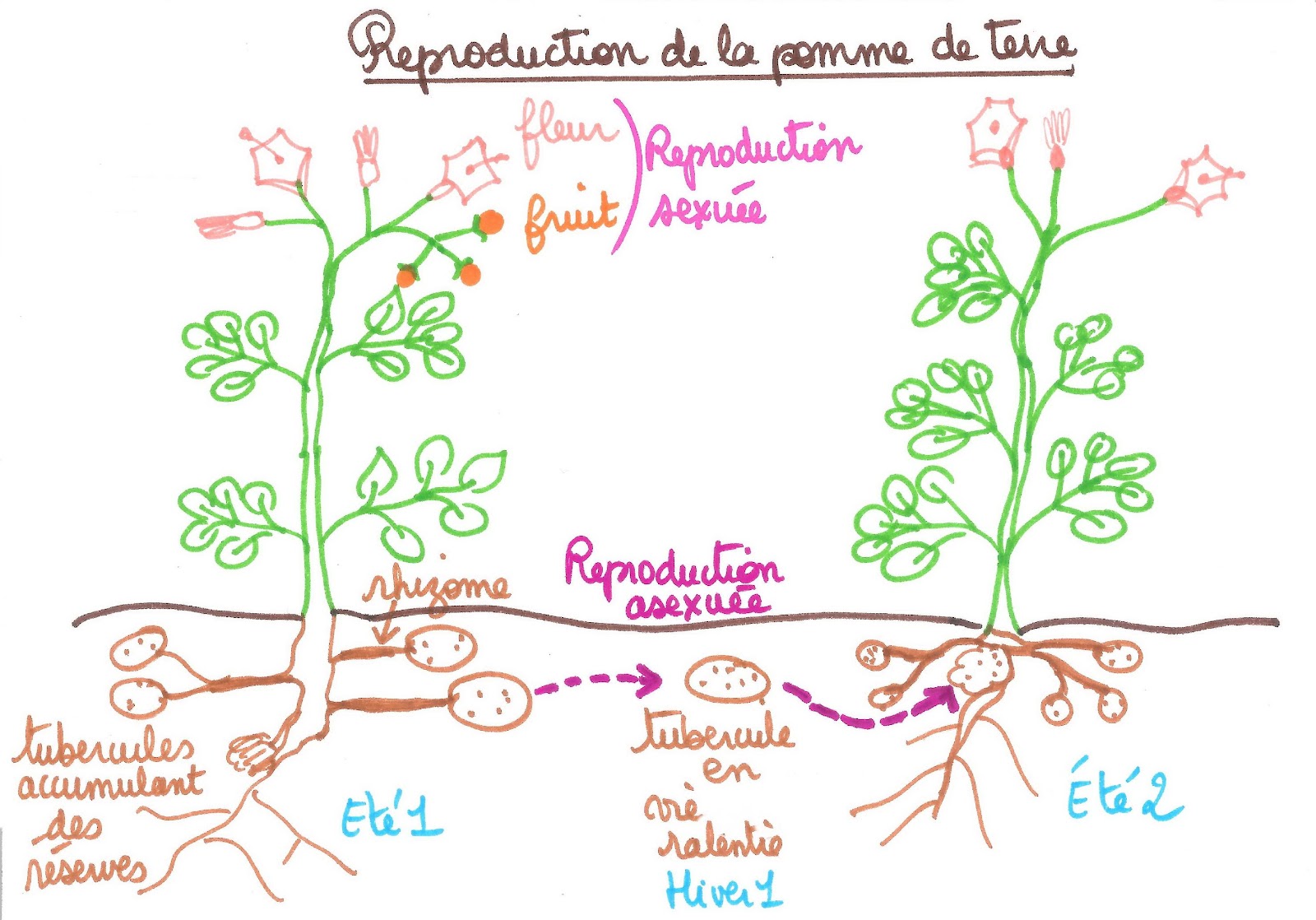
Concernant la reproduction asexuée ou peut citer par exemple la pomme de terre. C’est une plante vivace car à chaque printemps elle renaît de ses tubercules caulinaires restés sous terre. Si les tubercules sont dispersés, ils participent à la multiplication végétative, ou reproduction asexuée. En effet, chaque tubercule possède des bourgeons appelés « yeux ». Au printemps, le tubercule isolé développe de nouvelles tiges à l’origine d’un nouvel individu. Ces organes de réserve sont consommés car ils permettent un apport énergétique dans la durée : ils contiennent des réserves en sucre complexe, de l'amidon.
Document 10 : reproduction de la pomme de terre
©RS.2020
Document 11 : Pomme de terre germée et bourgeons de la pomme de terre
L’amidon est un polymère de glucose formé par une enzyme l’amylosynthase. Il se retrouve dans le stroma des chloroplastes sous forme de grains d’amidon appelés aussi granules. Il peut être stocké dans des plastes particuliers appelés « amyloplastes », organites très nombreux dans les organes de réserve comme le tubercule de pomme de terre. Chez de nombreuses espèces, l’amyloplaste ne possède qu’un seul gros grain d’amidon. Celui-ci se forme par accumulation de couches concentriques d’amidon et on peut facilement distinguer les stries de croissance. D’une plante à l’autre, les amyloplastes n’ont pas tout à fait la même forme. L’amidon est mis en évidence par le lugol de couleur brun-jaunâtre qui devient noir en sa présence.


Document 12 : Amyloplastes de pomme de terre colorés à l’eau iodée

©RS.2020
Concernant la reproduction sexuée, la graine, organe-puits, est à l’origine d’un nouvel individu et doit posséder des réserves permettant le développement de la plantule en attendant l’activation de la photosynthèse par les premières feuilles.
Les réserves de la graine sont généralement situées dans deux feuilles de réserves appelées cotylédons. Ces cotylédons peuvent être entourés d’un tissu de réserves particulier appelé albumen et l’on parlera dans ce cas de graine albuminée. Les graines ne possédant pas d’albumen sont appelées graines exalbuminées.
Document 13 : Structure d’une graine exalbuminée type haricot

Source : Dycotyledon seed diagram-fr.svg par LadyofHats / Traducteur Cehagenmerak via Wikimédia Commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Dycotyledon_seed_diagram-fr.svg
Certaines graines sont consommées pour leur apport énergétique durable : elles possèdent des réserves d'amidon. On parle de réserves amylacées, du nom de l'amylose, principale forme de polymérisation du glucose dans l'amidon. La réserve amylacée peut se trouver dans les cotylédons chez les graines exalbuminées comme le haricot ou dans l’albumen comme par exemple chez la graine de ricin où le caryopse de blé ou de maïs qui n’est autre qu’un fruit sec contenant une graine.
Document 14 : Caryopse de blé et nature de ses réserves

Source : Informations-nutritionnelles-grain-de-blé.svg
par Jkwchui, Jon C, via Wikimédia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Informations-nutritionnelles-grain-de-bl%C3%A9.svg
Les réserves des graines peuvent également être des réserves protéiques. On parlera ainsi de graines protéagineuses. Elles peuvent être mises en évidence par le réactif du biuret. Les protéines proviennent de l’assemblage des acides aminés produits lors de la photosynthèse. On peut citer en exemple la protéine « aleurone », souvent présente en grains dans les vacuoles des cellules. Quand les vacuoles sont très riches en aleurone, celles-ci se fractionnent en plusieurs petites vacuoles qui se déshydratent et on va observer des grains d’aleurone dans le cytoplasme des cellules qui ne sont rien d’autres que des amas de protéines cristallisées. Ceci s’observe très bien dans les cellules d’un cerneau de noix.
Document 15 : Coupe de cerneau de noix observé au microscope optique x100
L’aiguille montre des cristaux d’aleurone

©RS.2020
On sait aussi que certaines graines sont utilisées pour produire de l'huile comme les noix ou les amandes. On trouve dans les cellules de leur tissu de réserve des triglycérides non miscibles et formant des gouttelettes visibles dans le cytoplasme des cellules et appelées oléosomes. On parle de graines oléagineuses. On peut également en trouver dans le chloroplaste et dans des plastes spécialisés appelés oléoplastes. À l'écrasement d’un cerneau de noix sur une feuille de papier on remarque l'apparition d'une tâche translucide traduisant la présence de réserves oléagineuses.
Document 16 : Gouttelettes lipidiques observées au microscope optique x100 après écrasement d’une coupe de cerneau de noix

©RS.2020
Si de nombreux fruits sont consommés aussi bien par les animaux que par les humains, c'est parce qu'ils offrent également un apport énergétique. La saveur sucrée nous indique qu’on trouve des sucres simples dans leur tissu de réserve : glucose, fructose et leur forme associée, le saccharose. Ce dernier est formé grâce à une enzyme, la saccharose synthase. Ce sucre non réducteur, forme de transport par la sève, est présent chez de nombreux végétaux. Ayant une faible réactivité, il est peut-être stocké en quantité importante dans la vacuole de certains tissus comme chez la betterave sucrière ou la canne à sucre. La mise en évidence de sucres réducteurs peut se faire très facilement grâce à la liqueur de Fehling. Cette dernière se trouble à chaud en leur présence. L’accumulation de réserves au niveau du fruit facilite la dissémination des graines. Cet organe est donc toujours un organe-puits, il ne peut jamais devenir un organe-source pour la plante car le succès de sa dissémination dépendra entre autres de sa richesse en réserves.
Ainsi l’étude de la répartition des réserves montre que tous les organes de la plante peuvent être concernés par celles-ci. On remarque qu’il y a une dépendance réciproque des grandes fonctions métaboliques présentes dans la plante : la production de matière dépend non seulement de la photosynthèse mais également et surtout de la capacité de la plante à consommer et stocker les produits de celle-ci. De même, la libération de réserves sera corrélée à une diminution voire même une absence de la photosynthèse. Il y a une dépendance réciproque des deux phénomènes qui varient simultanément et qui sont fonctions l’un de l’autre. Comme ces corrélations sont relatives à la nutrition de la plante, on parle de corrélations trophiques.
Document 17 : Corrélations trophiques en été dans une plante herbacée

III Les molécules impliquées dans les interactions avec d’autres espèces.
On distingue deux catégories de molécules produites par les végétaux :
- les métabolites primaires directement impliqués dans la croissance, le développement et la reproduction d’un organisme ou d’une cellule. Ils sont souvent présents dans de nombreux organismes taxonomiquement éloignés. Ce sont les lipides, les glucides et les protides dont nous avons parlé précédemment.
- Les métabolites secondaires ne sont pas directement impliqués dans ces processus physiologiques fondamentaux. Ils possèdent par exemple une fonction écologique importante comme une fonction relationnelle. En général un métabolite secondaire est présent dans un ensemble taxonomique restreint d’organismes.
On peut citer comme premier exemple les tanins. Ce sont des polymères de phénol absents chez les animaux et que l’on trouve dans les racines, les feuilles, les tiges et les fruits.
Document 18: Molécule d’acide tannique
Source : Acide tannique.svg, par via via User_talk: Ronhjones, Wikimedia commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Tannic_acid.svg
Les tanins conféreraient une certaine protection aux végétaux les produisant. En effet, Les tanins associés aux mucoprotéines de la salive des herbivores donneraient des complexes moléculaires responsables d’une astringence et d’une sensation de sécheresse dans la bouche de ces derniers, et donc seraient responsables d’une action répulsive sur les animaux. Il a été démontré en 1981, que les arbres riches en tanins étaient moins broutés par les herbivores de la faune kényane que ceux qui en produisaient peu. Des mesures ont prouvé que les arbres augmentent leur production de tanins quand ils sont soumis à la prédation.
Document 19 : Grand Koudou

Source : kudu-753045_1920 ?????? Julien Legrand ??? Pixabay , https://pixabay.com/th/photos/kudu
Des tests en laboratoire ont montré par exemple chez les oies du Canada, que des aliments qui possèdent plus de 5 % de tanins comme l’acide férulique et l’acide tannique, sont clairement évités par ces oiseaux. Il a été prouvé que les tanins sont hépatotoxiques et que les animaux en consommant ont une croissance diminuée. L’ensemble de ces expériences montre le rôle protecteur des tanins vis-à-vis de la prédation.
Document 20 : Oie Bernache du Canada

Source : goose-1386066_1920, Image par jLasWilson de Pixabay
D’autres métabolites secondaires ont été localisés dans les vacuoles des cellules végétales ou chez les champignons, ce sont les Anthocyanes. On les retrouve aussi bien dans les pétales des fleurs qu’au niveau des fruits. Ils absorbent préférentiellement dans le jaune et le vert ce qui induit des couleurs bleues et rouges à l’observation. Les couleurs vives qu’ils induisent après absorption, attirent les insectes et les oiseaux pollinisateurs et favorisent ainsi la dispersion du pollen et des graines.
Document 21 : Spectre d’absorption d’un anthocyane, le malvidine 3O glucoside.
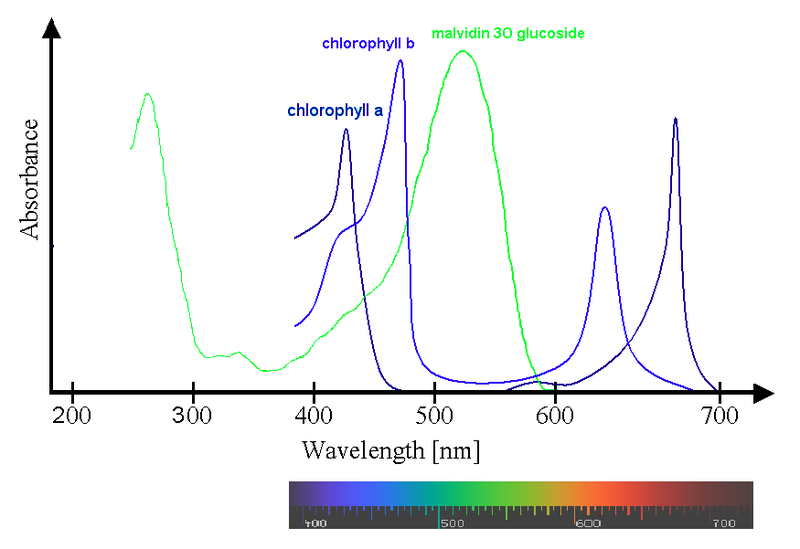
Source : Spectra Chlorophyll ab oenin (1) .PNG par “Pas avec » via Wikimedia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Spectra_Chlorophyll_ab_oenin_(1).PNG
Document 22 : Les fruits rouges sont riches en Anthocyanes

{kind=link}
Source : raspberries-4291303_1920 , Image par Thorsten Frenzel de Pixabay
Les principaux rôles physiologiques qui leurs sont attribués dans la plante sont, entre autres, l'absorption des radiations néfastes pour la chlorophylle b, le transport des monosaccharides, la régulation de la pression osmotique durant les périodes de sècheresse et de froid ou la régulation de la réponse antioxydative des plantes soumises à des facteurs de stress.
Devenir des produits de la photosynthèse - SVT - ENJEUX Term spé #5 - Mathrix
Date de dernière mise à jour : 08/06/2021