Le développement des plantes à fleurs
I Rappels et contexte d’étude :
Une plante à fleur est constituée d’un appareil végétatif comprenant des racines ancrées dans le sol et des tiges feuillées se développant en milieu aérien ou aquatique. On parle d’appareil racinaire et d’appareil caulinaire.
Document 1 : Organisation générale d’une plante à fleurs

Source : Schéma plante + fleur.png, par Fatlord , via Wikimedia commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Sch%C3%A9ma_plante_%2B_fleur.png
Chez les plantes à fleurs, l’embryon contenu dans la graine n’est qu’une ébauche de la future plante. Sa forme ne permet pas de définir les caractéristiques de celles-ci. Ce n’est que lors du développement de celle-ci que progressivement vont se mettre en place les caractéristiques de l’espèce.
Document 2 : Embryon d’une graine de Haricot

Source : Dycotyledon seed diagram-fr.svg par LadyofHats / Traducteur Cehagenmerak via Wikimédia Commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Dycotyledon_seed_diagram-fr.svg
On cherche à comprendre où est localisée à l’échelle du végétal la production des nouveaux organes ainsi qu’éclaircir les mécanismes cellulaires de cette production.
I Développement des racines
A/ Généralités
La racine est un des organes fondamentaux de la plante, permettant d’absorber l’eau et les sels minéraux nécessaires à la production de matière organique. Sa croissance est généralement souterraine.
Son extrémité perçoit les signaux en provenance de l’environnement capables de modifier son orientation. La racine aura donc une croissance orientée selon ceux-ci. On appelle géotropisme, la direction dans laquelle un être vivant se développe en relation avec la pesanteur. Il sera dit positif si la croissance se fait dans la même direction que la pesanteur et sera dit négatif si la croissance se fait à l’opposé de la direction de la pesanteur. On appelle thigmotropisme, la croissance orientée selon la présence d’obstacles et l’hydrotropisme, la croissance liée à l’humidité du sol
Document 3 : Les tropismes racinaires

©RS.2020
Sur une racine, on ne trouve ni bourgeons ni feuilles, c’est un organe non chlorophyllien.
Elle assure différentes fonctions qui sont :
• l’ancrage au sol
• l’absorption d’eau et de sels minéraux
• le stockage de réserves
• la sélection des ions
• la synthèse de certaines hormones végétales
La racine présente une symétrie axiale : elle est organisée en couches concentriques de cellules capables d’assurer différentes fonctions. Voici les différents tissus d’une racine :
- le rhizoderme : tissu superficiel de la racine, équivalent de l’épiderme de la feuille ou de la tige
- le cortex ou parenchyme, tissu de réserve qui assure le transport de l’eau et des sels minéraux absorbés
- l'endoderme dont la barrière subéreuse (cadre de Caspary) filtre les solutés de l’eau.
- le péricycle situé sous l’endoderme qui est à l’origine des racines latérales
- le phloème et le xylème, vaisseaux conducteurs séparés par une couche génératrice des cellules vasculaire appelée pro-cambium.
- la moelle ou parenchyme médullaire, au cœur du cylindre central qui peut avoir un rôle de réserve.
Document 4 : Structure d’une racine en coupe

Racine1.jpg, , Utilisateur: Cehagenmerak , image retravaillée d'après un schéma du Wikipédia chinois ., via Wikimédia Commons, Creative Commons Attribution-Share Alike 3.0 Unported , https://commons.wikimedia.org/wiki/File:Racine1.jpg
B/ Croissance de la racine
Si on trace sur une racine en croissance des traits équidistants à l'encre (Expérience de Sachs), on remarque rapidement que seules les marques situées vers l'extrémité de la racine s'écartent les unes des autres. La croissance est dite subterminale. C'est donc au niveau de l'apex qu'il faut chercher les sites responsables de la rhizogenèse (du grec rhiza =racine, génèse/fabrication des racines).
Document 5 : Expérience de marquage sur des racines de graines en germination.

©RS.2020
La pointe d'une racine comporte3 étages dans sa longueur, 3 zones révélant également un étagement dans la différenciation cellulaire.
- La zone de divisions séparée en trois parties :
- La coiffe : c'est le manchon cellulaire à l'extrémité de la racine (position terminale) qui a pour but de protéger le méristème racinaire. Les cellules les plus externes de cette coiffe sécrètent en abondance un mucilage avant de se détacher au contact du sol à cause des dommages mécaniques provoqués par le contact de la racine en croissance avec les particules du sol. La coiffe nécessite donc une régénération permanente de ses cellules, réalisée à partir de la zone d'entretien de la coiffe, du côté racinaire.
- Le centre quiescent : situé juste au-dessous de la coiffe, est la population cellulaire «souche». ll donne naissance, vers la partie distale (partie la plus éloignée du centre de la racine), aux cellules de la coiffe et, vers la partie proximale (partie la plus proche du centre de la racine), aux cellules méristématiques
- Le méristème racinaire : il contient des cellules non différenciées, présentant un gros noyau, sans vacuoles et assurant la production de nouvelles cellules par division (mitose). Il est divisé en trois méristèmes spécifiques.
- La zone d’élongation située à l’arrière du méristème, où les cellules nouvellement formées vont pouvoir s’allonger et amorcer une différenciation pour donner tous les tissus cités précédemment.
- La zone de différenciation où les différents tissus constitutifs de la racine se mettent en place et prennent leur fonction. Les cellules jusqu’alors indifférenciées, se différencient en acquérant un caractère spécifique. Le début de cette zone de différenciation sera occupé par la zone pilifère (riche en poils absorbants) qui finira par disparaître au fur et à mesure de l’épaississement du rhizoderme.
Document 6 : Coupe d’apex de racine d’ail observée au microscope

La zone méristèmatique se situant toujours à l’extrémité de la racine, l’apex, on parle de méristème apical racinaire (MAR). Elle est responsable de la croissance en longueur de la racine. Elle est divisée en trois méristèmes primaires, disposés en couches concentriques, chacun à l’origine d’un tissu primaire spécialisé et différencié :
- Le protoderme, situé le plus à l’extérieur de la zone méristématique, donnera le rhizoderme, c’est-à-dire le tissu de surface à l’origine des poils absorbants.
- Le Procambium ou méristème vasculaire, situé dans l’axe central de la racine, va donner une couche de cellules appelée cambium à l’origine des vaisseaux de xylème, de phloème et du péricycle.
- Le méristème cortical appelé aussi méristème fondamental, situé entre le protoderme et le procambium, donnera l’endoderme et le parenchyme cortical.
Document 7 : Structure et fonctionnement de l’apex d’une racine

©RS.2020
Chez certaines catégories de plantes à fleurs, peuvent apparaître des tissus secondaires permettant l’épaississement des organes. (hors programme)
II Développement des tiges
A/ Généralités
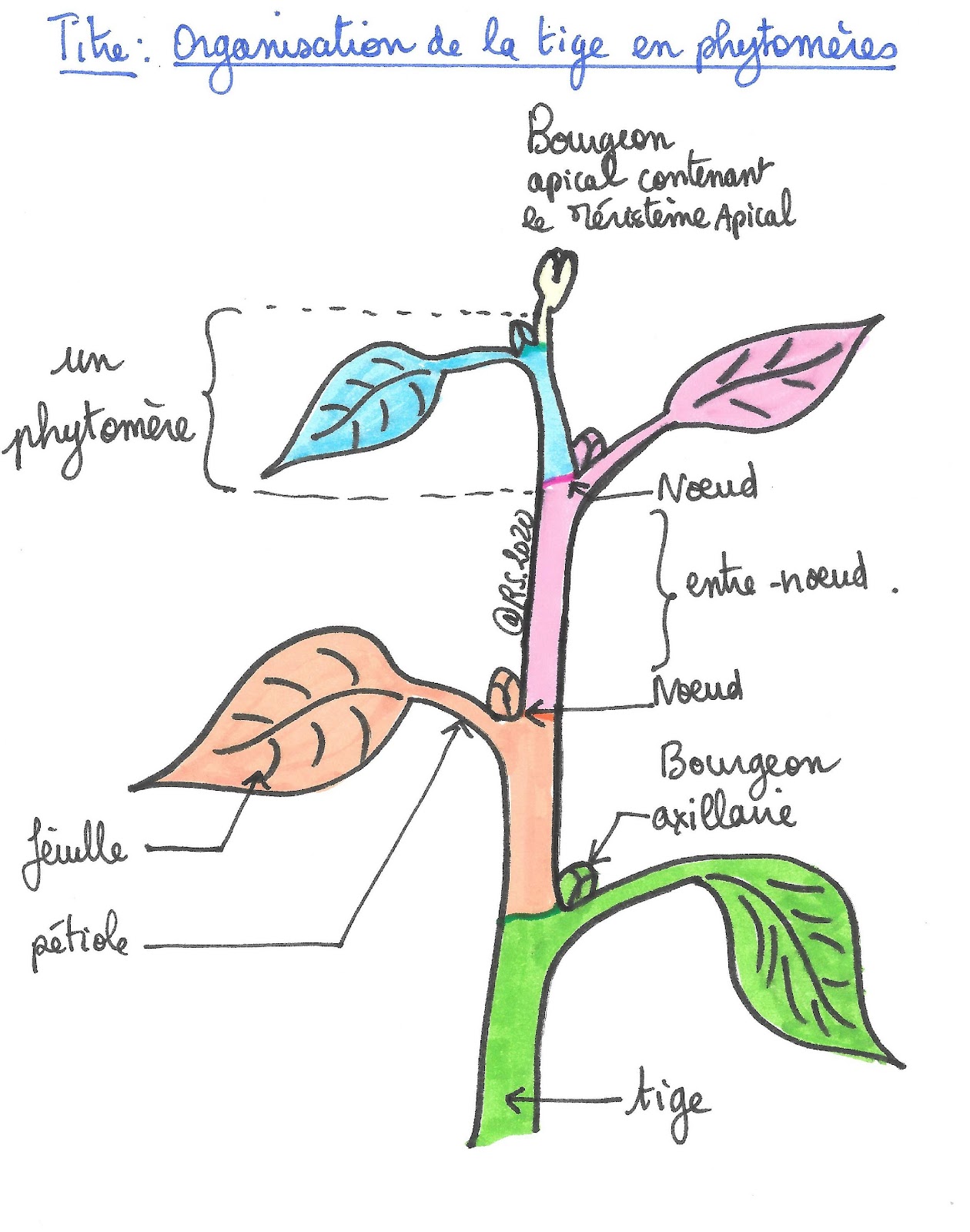
Qu’elle soit aérienne ou souterraine, qu’elle soit dressée ou rampante, une tige est constituée d’une succession de nœuds et d’entre-nœuds. Elle porte les feuilles et les organes reproducteurs. C’est également le lieu de transit des sèves brute et élaborée vers les organes.
Chaque section située entre 2 nœuds correspond à un module répétitif appelé « phytomère ». Chaque phytomère présente un départ régulier de feuilles. Le phytomère se met en place à partir du bourgeon terminal. On peut donc dire que ce dernier contrôle la croissance et l’organisation de la tige. Quand la plante peut se ramifier,on observe à l’aisselle des feuilles, un bourgeon axillaire. Ce dernier développe un rameau organisé lui aussi en phytomère, sous le contrôle du bourgeon terminal du rameau.
Document 8 : Organisation d’une tige en phytomères
L’observation d’une coupe de tige entre les noeuds montre l’organisation concentrique de cette tige en plusieurs couches tissulaires :
- L’épiderme possédant des stomates et qui est recouvert d’une fine cuticule, participe à la protection de la plante vis-à-vis de la déshydratation et permet de canaliser les flux de gaz par les stomates.
- Le parenchyme cortical permet la photosynthèse quand les tiges sont herbacées et jeunes avec un épiderme transparent.
- Le phloème et le xylème organisés en faisceaux criblo-vasculaires, assurent la conduction des sèves au travers de la plante.
- Le parenchyme médullaire situé entre les faisceaux peut servir pour la mise en réserve ou alors disparaître au profit d’une lacune
- le collenchyme parfois présent, est un tissu de soutien dont les cellules ont une paroi cellulosique très épaisse.
Document 9 : Structure d’une tige vue en coupe

B/ Croissance de la tige
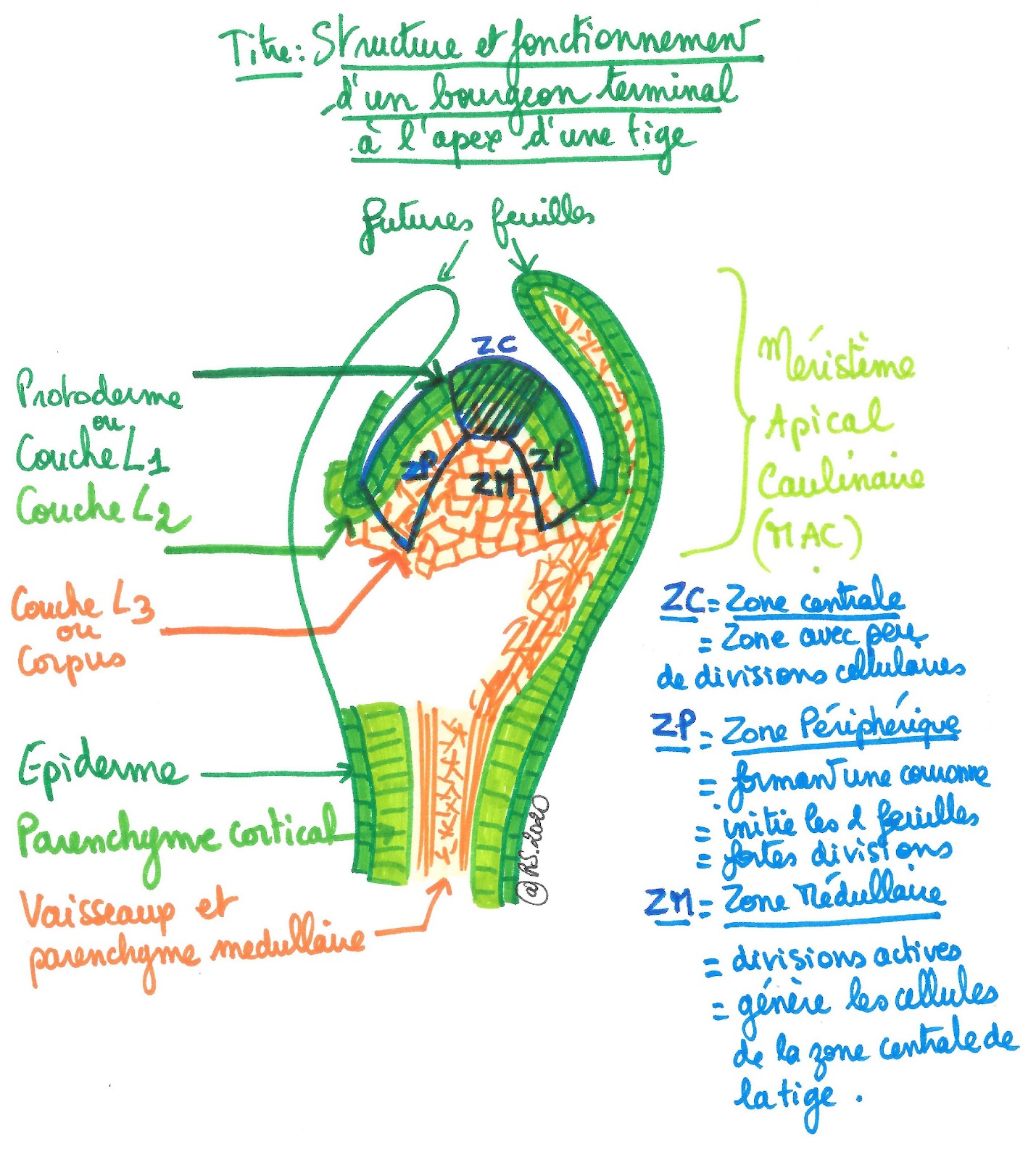
Les cellules de ces tissus, comme dans la racine, dérivent de cellules produites par méristème apical caulinaire ou MAC. Comme pour la racine, on distingue des tissus primaires dont la disposition est à l’origine de la structure primaire de la tige. Le méristème apical caulinaire (de la tige) est donc un méristème primaire qui permet la croissance en longueur de la tige.
Le méristème apical caulinaire se situe entre les deux futures feuilles du bourgeon. Il est constitué de trois épaisseurs concentriques de cellules, chacune à l’origine d’un tissu spécifique de la tige :
- la couche L1 ou protoderme, couches de cellules organisées, donnera, après différenciation, l’épiderme de la tige.
- la couche L2, couche de cellules également organisées, donnera le parenchyme cortical.
- la couche L3 ou corpus constituée de cellules non organisées, donnera l’ensemble des vaisseaux et du parenchyme médullaire de la tige.
L’activité mitotique n’est pas régulière sur l’ensemble du méristème apical.
- La zone centrale (ZC) présente peu de divisions cellulaires. Elle intervient peu dans l’organogenèse de la tige (mise en place des organes). Si besoin, elle fournit des cellules aux zones voisines.
- La zone périphérique (ZP) formant une couronne autour de la zone centrale, subit de fortes divisions et initie les deux feuilles.
- La zone médullaire (ZM) générera les cellules de la zone centrale de la tige.
Document 10 : Structure et fonctionnement d’un méristème apical caulinaire
Comme pour la racine, chez certaines plantes, des méristèmes secondaires peuvent apparaître et permettre une croissance en épaisseur ( pour donner un tronc par exemple). Si la plante est mature, le méristème apical peut se transformer en méristème floral.
III Un développement sous influence hormonale
Le développement d’un végétal correspond à la fois à l’augmentation de taille (la croissance) et à la formation de ses organes (organogenèse).
A/ Influence hormonale sur la croissance
1) Mise en évidence d’un messager chimique
La croissance d’une plantule à la sortie d’une graine est soumise à l’action de la lumière. La plantule pousse rapidement juqu’à sortir de terre pour atteindre la lumière. Une fois la surface atteinte, la croissance ralentit et la plantule jusqu’alors restée jaune, devient verte.
Document 11 : La croissance de plantules de lentilles à l’obscurité
Pour comprendre la cascade d’événements qui interviendraient dans une plante sous l’influence de la lumière, plusieurs scientifiques ont réalisé des expérimentations sur des coléoptiles de graminées qu’ils ont soumis à différents traitements.

Darwin fut le premier en 1880 à étudier le phénomène de la courbure phototropique du coléoptile d’avoine. Le coléoptile est un organe transitoire lors de la germination : il forme une gaine protectrice autour des pousses émergentes notamment chez les graminées.
Document 12 : Coléoptile à droite et sortie de la tige de maïs à gauche

Source : Mais kiemplant.jpg, par . Rasbak (Overleg). via wikimédia commons, CC-BY-SA-3.0-migré
https://commons.wikimedia.org/wiki/File:Mais_kiemplant.jpg
Lors de la germination le coléoptile est capable d’orienter sa croissance en direction de la lumière. On parle de phototropisme. Darwin a pu observer qu’en plaçant une plantule dans une boîte fermée et en perçant celle-ci d’un petit trou pour laisser entrer une lumière unidirectionnelle, le coléoptile se courbait vers cette source de lumière. Cette courbure résulte d’une différence de croissance entre la face exposée est la phase non exposée : cette dernière en grandissant plus vite induit une courbure du coléoptile du côté éclairé. En coupant le sommet du coléoptile et en reprenant les mêmes conditions expérimentales, Darwin remarqua une absence de courbure. Le résultat fut le même quand il plaça un cache sur l’apex du coléoptile. Il en conclut que l’apex du coléoptile contrôlait la croissance de ce dernier.
Document 13 : Expérimentation de Darwin

©RS.2020
Boysen-Jensen entre 1910 et 1913, chercha à identifier la nature du message responsable de la croissance de la tige en positionnant entre celle-ci et son coléoptile un bloc de gélose. Il observa la courbure du coléoptile. Il en conclut que le message contrôlant la croissance de la tige passait du coléoptile à celle-ci à travers le bloc de gélose. Söding, en 1923, testa d’autres éléments comme le mica (minéral isolant) , le platine (métal conducteur) et du beurre de cacao. Il remarqua à chaque fois une absence de courbure. Il en conclut que le message n’était pas de nature électrique, que c’était une substance non pas liposoluble mais plutôt hydrosoluble.
Document 14 : Expérimentation de Boysen-Jensen

©RS.2020
En 1931, Kogl et Haagen Smit identifient la structure chimique du messager qui porte le nom d'auxine (acide 3 indole acétique ou AIA). Il possède une chaîne acétique et un noyau indole.
Document 15 : Molécule d’auxine.
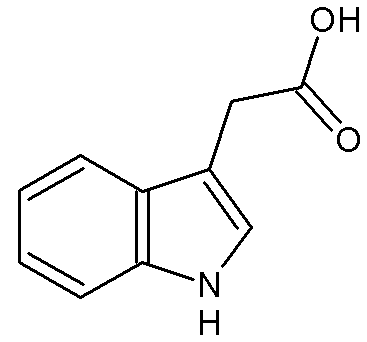
Source : IAAII.png par socrtwo,via Wikimédia commons, Domaine publique, https://commons.wikimedia.org/wiki/File:IAAII.png
2) L’action de l’auxine
Des expérimentations très simples peuvent être faites pour vérifier l’action de l’auxine sur la croissance des coléoptiles.
Sur un lot de graines ayant germé simultanément, différents tests sont effectués.
Première expérimentation :
- Deux graines vont servir de témoins : aucune intervention n’y sera effectuée.
- Deux graines suivantes (test n°1) vont subir une section de l’apex du coléoptile. L’apex étant le seul élément manquant dans le test, en fonction du résultat obtenu, le rôle de celui-ci dans la croissance sera ainsi prouvé ou pas.
Premiers résultats : après 48 heures,on constate une absence de croissance des coléoptiles dont l’apex a été sectionné (test n°1). On en conclut que celui-ci libère une substance responsable de la croissance.
Deuxième expérimentation : Pour confirmer que la substance soit bien de l’auxine, il suffit de réitérer la section de l’apex sur 2 graines germées (test n°2), et de déposer au niveau de la section, un cube de gélose contenant de l’auxine. Afin d’éliminer une quelconque influence de la gélose, il faut réaliser un témoin n°2 consistant à décapiter un coléoptile et à positionner un cube de gélose sans auxine sur la section. Ainsi par comparaison entre le témoin n°2 et le test n°2, il est possible de déterminer l’action de l’auxine seule.
Deuxième résultat : voir document ci-dessous
Document 16 : Expérimentation de section d’apex de coléoptiles de graminée et dépôt d’auxine.

Exploitation des données :
Pour confirmer le rôle de l’auxine, il faut calculer le taux d’accroissement des coléoptiles. En effet, il faut comparer ce qui est comparable. Si les coléoptiles avaient eu exactement la même taille au démarrage de l’expérience, on aurait pu simplement comparer leur taille 48 heures après. Cependant ce n’est pas le cas. On le voit très bien par exemple sur le coléoptile n°1 : c'est le plus grand coléoptile à la fin de l'expérience mais aussi au départ ! Il a peut-être proportionnellement moins grandi que les n°5, 6, 7 ou 8. En rapportant la taille mesurée au bout de 48h à la taille mesurée au moment de la décapitation, on obtient un taux de croissance pour chaque coléoptile. On pourra ainsi comparer les taux respectifs de chacun et dire lequel a subi la plus forte croissance et ainsi conclure sur le rôle de l'auxine.
Document 17 : Pourcentage de croissance des différents coléoptiles testés

Les résultats au bout de 48 heures montrent que les coléoptiles disposant d’un cube de gélose avec de l’auxine présentent un taux de croissance de l’ordre de 350 % contre 180 % pour ceux dont l’apex a été sectionné et qui n’ont reçu que de la gélose. Les coléoptiles dont l’apex n’a pas du tout été sectionné ont atteint un pourcentage de croissance d’environ 260 %.
Interprétation :
On peut donc affirmer, suite à cette expérience, que l’auxine est donc bien un facteur de croissance produit par l’apex et migrant vers les zones situées en dessous de celui-ci. On parle de transport vertical en direction des racines ou transport basipète.
Document 18 : Transfert de l’auxine de l’apex vers la tige
©RS.2020

Ce facteur de croissance possède une double action sur l’élongation cellulaire au niveau de la zone d’élongation subapicale :
- une action à court terme sur la plasticité pariétale (plasticité de la paroi). En effet, les cellules végétales sont entourées d’une paroi protectrice rigide constituée de cellulose. Celle-ci peut être un frein à la croissance cellulaire dans les zones d’élongation proche des zones méristèmatiques. L’auxine va provoquer le relâchement de la paroi permettant ainsi à la cellule de grandir.
- une action à long terme sur l’expression de gènes participant au métabolisme de la croissance : elle régule la synthèse d’ARNm codant pour des protéines nécessaires à l’élongation. Une électrophorèse permet de mettre en évidence dans la tige en croissance, à la fois les protéines de croissance et l’auxine.
B/ Influence hormonale sur l’organogénèse
1) Les gibbérellines
Les gibbérellines sont des hormones végétalesproduites au niveau des méristèmes des bourgeons terminaux racinaires et caulinaires, des jeunes feuilles de l’embryon. Elles permettent l’allongement des cellules des entrenoeuds ainsi que le débourrement des bourgeons. Elles stimulent la croissance du fruit et retardent sa maturité. Elles n’ont pas d’action connue sur les racines.
Document 19 : Molécule de gibbéréline GA452DII, la plus connue.
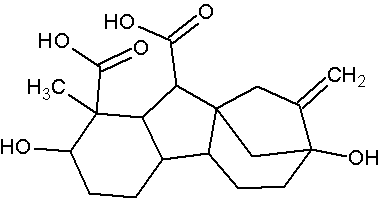
Source : GA452DII.pngpar socrtwo,via Wikimédia commons, Domaine publique, https://commons.wikimedia.org/wiki/File:GA452DII.png
2) Les cytokinines
Les cytokinines sont des composés proches des bases puriques.
Document 20 : Molécule d’une cytokinine, ici la zéatine
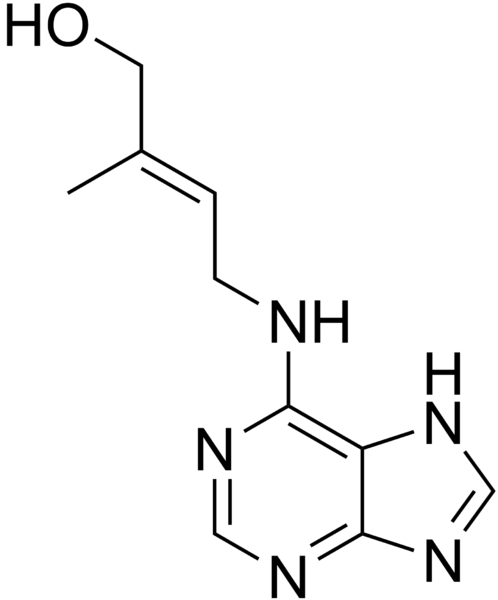
Source : Zeatin.png par Edgar181 via Wikimédia Commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Zeatin.png
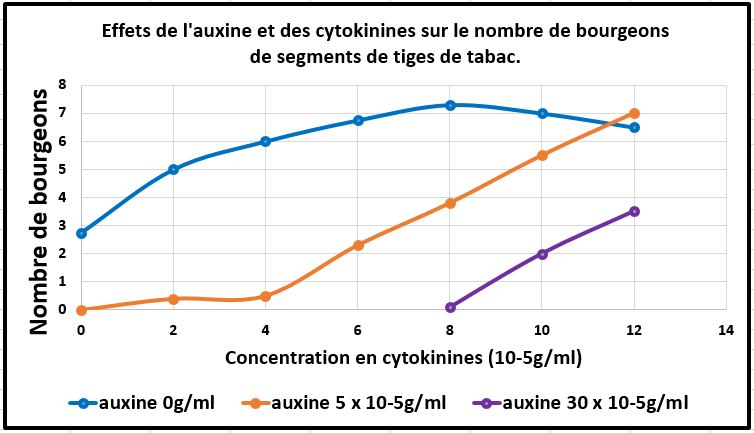
Elles sont fabriquées au niveau de l’apex racinaire et sont transportées verticalement en direction de la tige : on parle de transport acropète. Elles induisent la différenciation des bourgeons sous réserve de la présence de faibles doses d’auxine. En effet des concentrations élevées d’auxine versées sur des segments de tabac en culture, inhibent la différenciation des bourgeons (courbes orange et violette sur le graphique ci-dessous) alors qu’en l’absence d’auxine, les cytokinines induisent une forte production de bourgeons (courbe bleue ci-dessous). Les cytokinines ont des propriétés activatrices de la division cellulaire et sont impliquées dans la différenciation et donc dans la morphogenèse.
Document 21 : Effets de l'auxine et des cytokinines sur le nombre de bourgeons de segments de tiges de tabac par rapport aux taux des deux hormones végétales. d’après les données de Miller et Skoog, 1953
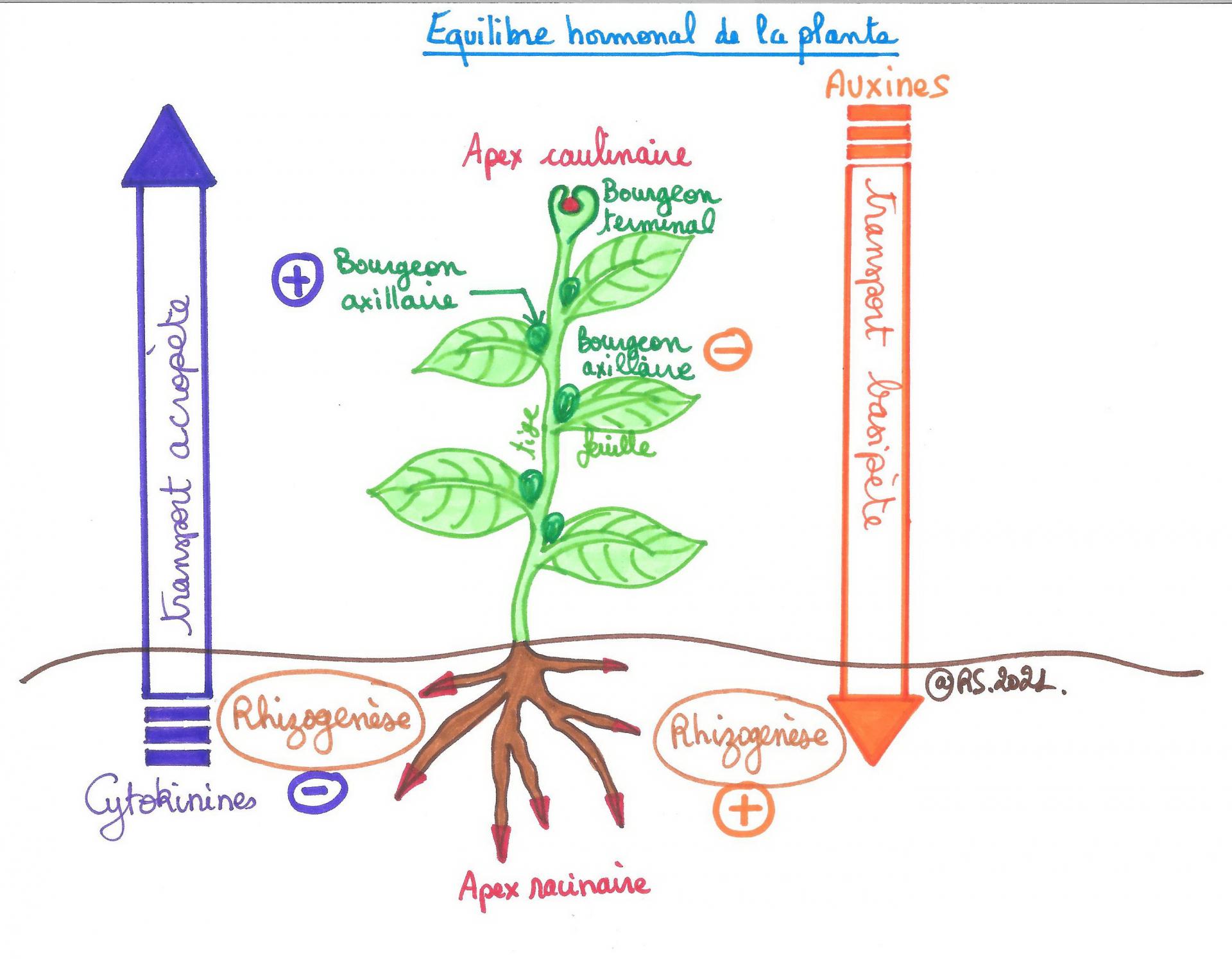
Ainsi à l’échelle de la plante plusieurs hormones sont produites et chacune agit spécifiquement :
- les auxines sont produites au niveau des bourgeons apicaux et favorisent l’enracinement tout en s’opposant au développement des bourgeons axillaires. On parle de dominance apicale.
- les cytokinines sont produites principalement au niveau des racines et favorisent le développement des bourgeons axillaires. Plus les bourgeons axillaires sont éloignés de l’apex caulinaire plus ils peuvent se développer et des ramifications apparaissent.
L’organogenèse dépendra donc des proportions relatives de ces deux hormones. L’organogenèse est une affaire d’équilibre hormonal.
Document 22 : Un équilibre hormonal chez la plante à fleurs
©RS.2020
Les cellules végétales sont totipotentes : elles peuvent se différencier différemment selon les catégories d’hormones reçues et selon la concentation de celles-ci. Grâce à la propriété de totipotence des cellules végétales, il est ainsi possible d’orienter l’organogenèse en jouant sur la balance hormonale dans une culture in vitro. Ainsi une forte concentration d’auxines par rapport à celle de cytokinines stimule la formation de racines ou rhizogénèse tandis qu’un rapport faible entraîne au contraire la caulogenèse c'est-à-dire la formation de tiges
.C/ L’influence de l’environnement
Le développement de la plante contrôlé par des hormones végétales peut subir influence de l’environnement
On peut citer l’exemple de ces arbres soumis à des vents réguliers et dont la croissance de l’appareil caulinaire ne sera pas vertical. Celui-ci adoptera un port dit en drapeau.
Document 23 : Port en drapeau d’un arbre

Source : tree-510047_1920 par LoggaWiggler de Pixabay , https://pixabay.com/fr/photos/arbre-tortueux-vent-pli%C3%A9-noueux-510047/
La température peut également modifier la croissance des plantes vivaces. En effet, à l’automne les arbres perdent leurs feuilles car celles-ci sont sensibles au gel. Ils vont ainsi entrer en vie ralentie : les bourgeons vont rester en dormance pendant l’hiver. Au printemps, le retour des températures douces et d’une luminosité plus importante et plus longue dans la journée, active la production des hormones végétales à l’origine du débourrement des bourgeons et de la croissance des rameaux.
Document 24 : Débourrement d’un bourgeon au printemps

Source : bud-1347462_1920, Image par Jiricek72 de Pixabay, https://pixabay.com/fr/photos/bourgeon-printemps-fermer-arbre-1347462/
La croissance des plantes à fleurs est soumise à l’action de la lumière : on parle de phototropisme. On sait que la croissance cellulaire et sous le contrôle de l’auxine. Il a été remarqué que le côté éclairé de la tige présente une courbure orientée vers la lumière résultant d’une inégalité de croissance entre le côté éclairé et le côté non éclairé, ce dernier ayant une croissance plus importante. L’étude de la répartition de l’auxine dans une tige présentant une courbure montre que la concentration en cette hormone est plus importante du côté non éclairé que du côté éclairé, sans pour autant qu’on sache encore l'expliquer.
Document 25 : Exemple de développement lié à un phototropisme .
La plante à l’abri de la lumière derrière la commode, oriente ses tiges vers la fenêtre tout en contournant l’obstacle.

©RS.2020
Le végétal perçoit également la gravité, force qui s’exerce sur tous les objets terrestres, ce qui permet d’orienter sa croissance. La plante peut percevoir assez rapidement, en moins d’une minute, un changement d’orientation, par exemple dans le cas où le vent ou un animal déplace une de ses branches. Ne disposant pas de système nerveux comme nous, la réponse de la plante va mettre plusieurs minutes pour s’effectuer. La coiffe possède dans sa partie centrale un ensemble de cellules allongées formant le columelle. Ces cellules appelées statocytes contiennent des organites appelés statolithes, permettant la perception de la gravité. Ce sont en général de gros amyloplastes qui sédimentent sous leur propre poids ou parfois des cristaux de sulfate de baryum.
Document 26 : Réorientation de la racine par Gravitropisme

{kind=link}
Développement des plantes à fleurs- SVT - ENJEUX Term spé #3 - Mathrix
Date de dernière mise à jour : 31/08/2021