La cellule
La notion de cellule fut construite entre le xviiieme siècle et le xixeme siècle grâce à la découverte et au perfectionnement des microscopes. La théorie cellulaire est admise par la communauté scientifique à la fin du xixeme siècle.
I La cellule, découverte et débats
En 1665, Robert Hooke (1635-1703), scientifique anglais, observe pour la première fois au microscope optique qu’il a mis au point, un tissu d’un être vivant, celui du liège. Il y observe des petites chambres régulières qu’il appelle alors « cellules » du latin « cellula » qui veut dire « petite chambre ». Cependant ces cellules sont mortes : en effet ce ne sont que les restes des parois cellulosiques de celles-ci mais il ne le savait.
Le fonctionnement du microscope de Hooke était simple : il a assemblé deux lentilles convexes, un oculaire et un objectif, permettant ainsi de grossir les objets jusqu’à 30 fois. Les lentilles de l’époque présentaient des défauts optiques ce qui nuisait à la qualité de l’image. La lumière nécessaire au fonctionnement du microscope était fournie par une lampe à huile dont les rayons concentrés par un ballon à eau éclairaient l’objet à observer. Les rayons réfléchis remontaient dans le tube optique traversant les lentilles, formant ainsi une image.
À la même époque, d'autres observations sont réalisées grâce à des microscopes optiques : globules rouges, spermatozoïdes, bactéries, mais la cellule n’était pas encore reconnue comme l’unité de base du vivant car n’était pas observable tout le temps.
Microscope inventé par Robert Hooke en 1665 et dessin de ses premières observations de cellules.
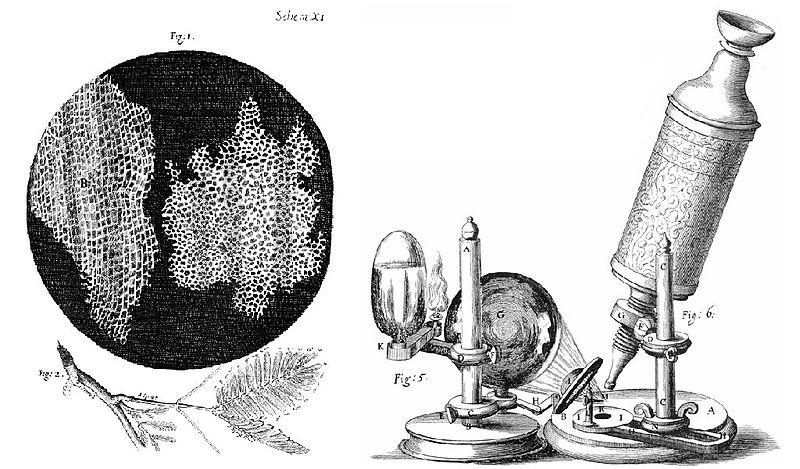
Sources : Hooke-Microscope-cork.jpg, source : Micrographie, Robert Hooke (1635-1702). Imagen montada y subida por Alejandro Porto ., domaine public, via wikimédia commons, https://commons.wikimedia.org/wiki/File:Hooke-Microscope-cork.jpg
Malgré l’existence du microscope, pendant très longtemps la théorie fibrillaire resta la théorie reconnue par les savants de l’époque. En effet la simple dissection des organismes vivants mettait en évidence la présence de nombreuses fibres aussi bien chez les animaux que chez les végétaux. Il était donc admis que la fibre était l’élément constitutif du vivant. De plus, pendant longtemps de nombreux scientifiques n’ont pas reconnu l’importance des apports de la microscopie pour la compréhension du vivant.
C’est dans les années 1830, que grâce à de nombreuses études microscopiques portant sur différents tissus végétaux, que le scientifique écossais Robert Brown (1773-1858) découvrit et décrit un élément constitutif important des cellules : le noyau. C’est le scientifique allemand Théodore Schwann (1810-1882) qui en réalisant de nombreuses études microscopiques du vivant, finit par établir la correspondance entre les cellules animales et végétales : elle possédait une organisation commune. Ainsi en 1838, Schawnn fut le premier scientifique à énoncer la théorie cellulaire du vivant : tous les tissus, animaux et végétaux, sont constitués de cellules contenant un noyau. La cellule dans cette théorie est considérée comme l'unité structurelle et fonctionnelle du vivant. Les propriétés des tissus apparaissent alors comme résultant des caractéristiques des cellules les constituant. Cette théorie est cependant rejetée par de nombreux scientifiques.
En 1855 plusieurs travaux ont mis à jour la régénérescence des tissus par division des cellules. Ceci induit une acceptation définitive de la théorie cellulaire indiquant que la cellule est l’unité structurale et fonctionnelle de tous les êtres vivants et que toute nouvelle cellule se forme à partir d’une cellule préexistante par division de celle-ci. La cellule est effectivement observée chez tous les êtres vivants : animaux, végétaux, mais aussi les champignons et les bactéries.
La découverte de l’unité cellulaire est donc liée à l’invention du microscope. Plus récemment l’invention du microscope électronique a permis l’exploration de l’intérieur de la cellule et la compréhension du lien entre échelle moléculaire et cellulaire.
II Le modèle cellulaire
La cellule est un espace séparé de l’extérieur par une membrane plasmique. Elle contient un liquide appelé cytoplasme. C’est un système constitué de molécules et qui est le siège d’un métabolisme permanent. On appelle métabolisme cellulaire, l’ensemble des réactions chimiques qui se déroulent à l’intérieur d’une cellule et qui lui permettent notamment de se maintenir en vie, de grandir, de se multiplier ou encore de répondre aux différents stimuli qu’elle peut recevoir.
L’information génétique à l’origine de la structure de la cellule et de son métabolisme, se présente sous la forme d’acide désoxyribonucléique ou ADN. Chez les organismes eucaryotes (animaux, végétaux, champignons) l’information est contenue dans le noyau, un des organites intervenant dans le métabolisme complexe de ceux-ci. Chez les organismes procaryotes (bactéries, archées), l’ADN est directement présent dans le cytoplasme.
La description des différents organites présents dans la cellule s’est nettement améliorée dans les années 1960 grâce à l’invention du microscope électronique. Dans un microscope optique l’image est créée par une absorption différentielle de la lumière par le tissu.
Dans un microscope électronique ce n’est pas un faisceau de photons qui est émis mais un faisceau d’électrons à haute tension. L’orientation du faisceau d’électrons transmis par l’échantillon grâce à des lentilles magnétiques sur un écran phosphorescent permet d’obtenir une image optique de celui-ci. On parle de microscope électronique à transmission ou MET. L’image obtenue est en noir et blanc et porte le nom d’électronographie.
Electronographie de cellule animale:

Cellule animale au microscope électronique à transmission : microscopie électronique.jpg par Itayba sur Wikipédia hébreu, via Wikimédia Commons, CC-BY-SA-3.0 https://commons.wikimedia.org/wiki/File:Electron_microscopy.jpg
La microscopie électronique à balayage ou MEB permet d’obtenir des images en haute résolution de la surface d’un échantillon. En effet la surface de l’échantillon recevant le faisceau d’électrons réémet certaines particules qui vont être analysées par différents détecteurs. Ces derniers peuvent reconstruire une image en trois dimensions de la surface.
Fourmis observée au MEB:

Ant SEM.jpg
Par Gouvernement des États-Unis via Wikimédia Commons, Domaine public, https://commons.wikimedia.org/wiki/File:Ant_SEM.jpg
Ainsi la microscopie électronique a permis de décrire précisément les différents organites présents dans les cellules eucaryotes. Ceci a permis d’établir du lien entre les découvertes de biochimie, de génétique, de biologie moléculaire et de comprendre les différentes fonctions cellulaires. Ainsi par le décryptage des mécanismes moléculaires du fonctionnement cellulaire, le lien a été établi entre l’échelle moléculaire et l’échelle cellulaire.
Structure d’une cellule procaryote :

Source : Diagramme d une cellule procaryote.jpg par Mariana Ruiz [user:LadyofHats] Trduit par Baptiste Deleplacevia Wikimedia commons, Domaine public, https://commons.wikimedia.org/wiki/File:Diagramme_d_une_cellule_procaryote.jpg?uselang=fr
Structure d’une cellule eucaryote animale :

Traduction :
Nucleo : noyau
Nucleolo : nucléole
Ribosomi : ribosome
Reticulo endopamatico rugoso et liscio : réticulum endoplasmique rugueux et lisse
Apparato de golgi : appareil de Golgi
Mitocondri : mitochondrie
Centrioli : centriole
Citoplasma : cytoplasme
Lisosoma : lysosome
Vescicola : vésicule
Citoscheletro : cytosquelette
Source : Struttura della cellula animale.svg, source propre montage d'œuvres originales de MesserWoland et Szczepan1990 , créé avec Inkscape, basé sur les graphiques de en wiki, utilisateur: Giac83, via Wikimédia Commons, CC-BY-SA-3.0-migréhttps://commons.wikimedia.org/wiki/File:Struttura_della_cellula_animale.svg
Structure d’une cellule eucaryote végétale:
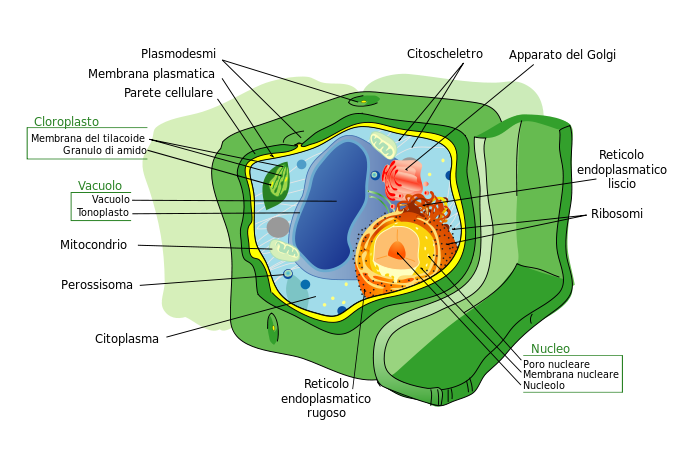
Traduction :
vacuolo : vacuole
membrana plasmatica : membrane plasmique
parete cellulare : paroi cellulaire
plasmodesmi : plasmodesmes (système de communication entre cellules permettant le passage de l’eau)
perossisoma : peroxysome (organite chargé de détoxifier la cellule)
Source : Struttura della cellula vegetale.svg par L'édition de Gia.cossa de l'original par Mariana Ruiz LadyofHats via wikimédia commons, domaine public, https://commons.wikimedia.org/wiki/File:Struttura_della_cellula_vegetale.svg

©RS.2019
III La membrane plasmique
La membrane plasmique appelée aussi membrane cellulaire, membrane cytoplasmique ou plasmalemme, est une membrane de 5 à 10 nm d'épaisseur séparant l’intérieur de la cellule, le cytoplasme, de l’extérieur ou milieu extracellulaire. La membrane a un rôle biologique important car elle isole la cellule de son environnement.
Elle est formée de phospholipides et de protéines. Les phospholipides sont des molécules organiques composées d'un groupement contenant un atome de phosphore (groupement hydrophile, ayant de l'affinité pour l'eau, c'est-à-dire lipophobe) et d'une partie lipidique (partie lipophile, ayant de l'affinité pour les lipides, c'est-à-dire hydrophobe).
La membrane correspond à une association de deux feuillets sombres de phospholipides épais de 2 nm chacun qui se font face par leur pôle hydrophobe : la partie hydrophile est orientée vers le milieu extracellulaire pour le feuillet externe et vers le milieu intracellulaire pour le feuillet interne tandis que la partie lipophile ou hydrophobe est orientée vers l'intérieur de la membrane. Ces deux feuillets sont séparés par un feuillet plus clair épais de 3,5 nm qui correspond à l'espace entre les deux parties hydrophobes de chaque couche de phospholipides.
Entre les molécules de phospholipides se trouvent des protéines dites intrinsèques et du cholestérol (uniquement chez les cellules animales). La membrane plasmique forme un milieu fluide où les lipides et les protéines migrent latéralement.
Sur les faces externe et interne constituées par les pôles hydrophiles des phospholipides membranaires, se fixent des protéines de surface intrinsèques sur lesquelles s’accrochent le cytosquelette côté interne et la matrice extracellulaire côté externe.
Hormis son rôle de protection, les différentes propriétés des membranes cellulaires, échanges de matière et d'information, sont essentiellement dues à la présence de protéines intrinsèques transmembranaires qui assurent différentes fonctions dont :
-
la perméabilité sélective avec par exemple les aquaporines laissant passer l’eau sans laisser passer les sels minéraux, les différents canaux ioniques,
-
la reconnaissance de signaux chimiques avec les récepteurs hormonaux,
-
la présentation de marqueurs antigéniques provenant d’agents pathogènes.
Au niveau du feuillet externe, des groupements glucidiques (carbohydrates) sont associés à des phospholipides et à des protéines. Certaines d’entre elles comme le glycocalyx, interviennent dans l’adhérence cellulaire (ensemble des mécanismes favorisant l’adhésion des cellules entre elles pour former un tissu).
La membrane peut être associée à des protéines dites extrinsèques (périphériques) localisées en dehors de la bicouche phospholipidique et sont soit ainsi entièrement extracellulaires soit entièrement intracellulaires. Elles interagissent avec la membrane par des liaisons électrostatiques de type liaisons hydrogènes au niveau de protéines intrinsèques. Elles interviennent alors dans le fonctionnement de ces dernières.
Structure de la membrane plasmique :

Source : Cell membrane detailed diagram 4-FR.svg derivative work: Dosto (d), Cell_membrane_detailed_diagram_4.svg: *derivative work: Dhatfield (talk), Cell_membrane_detailed_diagram_3.svg: *derivative work: Dhatfield (talk, Cell_membrane_detailed_diagram.svg: LadyofHats Mariana Ruiz, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Cell_membrane_detailed_diagram_4-FR.svg?uselang=fr
Certains organites comme les lysosomes, le réticulum endoplasmique, l’appareil de Golgi sont délimitées par une simple membrane alors que d’autres comme le noyau, les chloroplastes et les mitochondries le sont par une double membrane car issus d’une endosymbiose. La théorie endosymbiotique, ou hypothèse de l'endosymbiose, est l'hypothèse selon laquelle les chloroplastes et mitochondries des cellules eucaryotes, du fait qu’ils possèdent leur propre ADN et doués de division, proviennent de l’incorporation (endocytose) par certaines archées, des bactéries avec lesquelles elles auraient entretenu une relation endosymbiotique. Elle s'oppose à la théorie autogénique (ou hypothèse autogène) qui postule que le système endomembranaire (chloroplaste, mitochondrie et noyau), ainsi que le cytosquelette des cellules eucaryotes, ait évolué par complexification à partir d'une cellule procaryote ancestrale.
Théorie de l’endosymbiose :
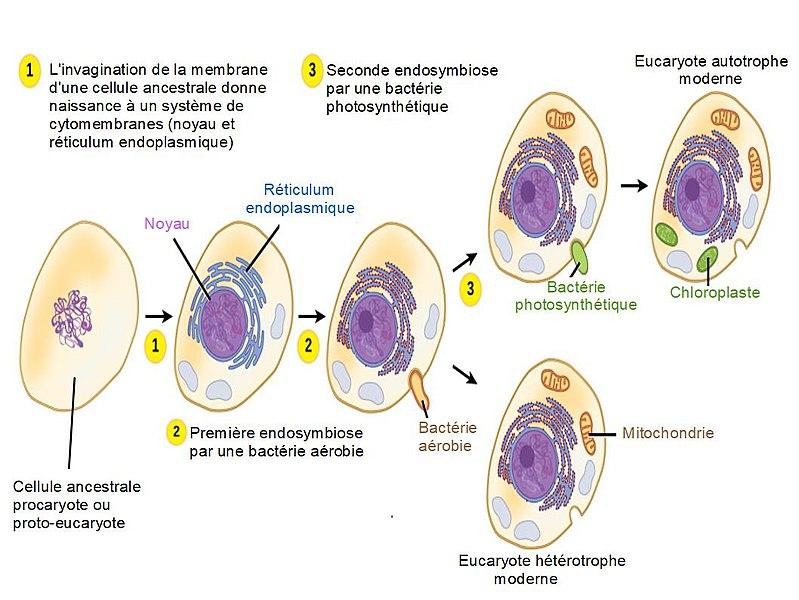
Source : Théorie endosymbiotique.jpg par Salsero35 via Wikimédia Commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Th%C3%A9orie_endosymbiotique.jpg
Origine des eucaryotes selon la théorie de l’endosymbiose:
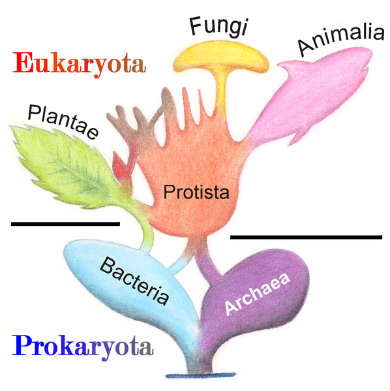
Source : Tree of Living Organisms 2.png par Maulucioni y Doridí via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Tree_of_Living_Organisms_2.png
Les archées, ou Archaea (du grec ancien ?ρχα?ος, « originel, primitif »), anciennement appelés archéobactéries, sont des microorganismes unicellulaires procaryotes, c'est-à-dire des êtres vivants constitués d'une cellule unique qui ne comprend ni noyau ni organites, à l'instar des bactéries. D'apparence souvent semblable à ces dernières, les archées ont longtemps été considérées comme des bactéries extrêmophiles particulières, jusqu'à ce que les recherches phylogénétiques sur les procaryotes, commencées en 1965, aboutissent, avec les travaux de Carl Woese et George E. Fox3, à la publication en 1977 d'un arbre phylogénétique fondé sur les séquences de certains gènes des organismes étudiés, arbre dans lequel les procaryotes étaient scindés en deux domaines distincts, celui des bactéries et celui des archées. Du point de vue de leur génétique, leur biochimie et leur biologie moléculaire, les archées sont des organismes aussi différents des bactéries que des eucaryotes. Les enzymes impliquées dans la lecture de l’ADN sont apparentées à celles des eucaryotes et non à celles des bactéries, de même que la présence de certaines protéines de structure des chromosomes (histones). Voilà pourquoi la théorie endosymbiotique est aujourd’hui mondialement reconnue et qu’il est admis que l'arbre des eucaryotes prend naissance parmi des archées.
Les archées ont longtemps été vues comme des organismes essentiellement extrêmophiles présents notamment dans les sources hydrothermales océaniques, les sources chaudes volcaniques ou encore les lacs salés, mais on en a découvert depuis dans tout une variété de biotopes qui ne sont pas nécessairement extrêmes, tels que le sol, l'eau de mer, des marécages, la flore intestinale et orale et même le nombril humain. Les archées seraient particulièrement nombreuses dans les océans, et celles faisant partie du plancton constitueraient l'un des groupes d'organismes les plus abondants de la Terre. Les archées interviennent par ailleurs de façon non négligeable dans le cycle du carbone et le cycle de l'azote. On ne connaît pas vraiment d'exemple d'archée pathogène ou parasite, mais elles sont souvent mutualistes ou commensales. Les archées méthanogènes de l'intestin humain et des ruminants participent ainsi favorablement à la digestion.
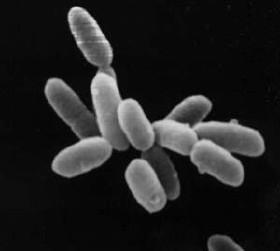
Archée :
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Source : Halobacteria.jpg, Par NASA, via Wikimédia Commons, domaine public, https://commons.wikimedia.org/wiki/File:Halobacteria.jpg
{kind=link}
Date de dernière mise à jour : 19/06/2021